Фаги
Фаги - вирусы бактерий, актиномицетов - имеют некоторые особенности, которые необходимо рассмотреть несколько подробнее.
Строение фага более сложно, чем строение вирусов животных и растений. Довольно своеобразна морфология фага. У него различают головку, имеющую овальную форму, иногда шестигранную, призматическую, иногда круглую. От головки отходит более или менее длинный полый отросток. Фаг сравнивают с барабанной палочкой, булавкой, головастиком. По своим размерам фаги относятся к средним по величине вирусам. Диаметр головки их составляет 60-90 ммк, длина отростка - 250 ммк, толщина - 10-25 ммк. Величина фагов довольно изменчива. Даже разные варианты (типы) одного и того же вида фага могут сильно различаться по своим размерам. Молекулярный вес фага 200 млн.
Частица фага является нуклепротеидом и состоит из белка (50-60%) и ДНК (45-50%). Некоторые фаги содержат небольшое количество липоидов (1,5-2%). Белок образует оболочку фага, а ДНК находится во внутреннем пространстве головки фага. Белковая оболочка состоит из большого числа белковых частиц, называемых субъединицами.
Некоторые ученые считают, что ДНК фага имеет очень небольшую примесь белка, который отличается от белковой оболочки фага. ДНК различных фагов отличаются друг от друга и от ДНК клеток бактерий, в которых они обитают. Кишечная палочка содержит ДНК около 5%, а лизирующие ее фаги - около 45%. Кроме того, ДНК кишечной палочки имеет основание цитозин, а ее фаг содержит другое основание, несколько отличающееся от цитозина.
Фаги являются паразитами бактерий (фаг - пожиратель). Они найдены почти у всех болезнетворных для человека и животных бактерий и у многих непатогенных микробов, в том числе у молочнокислых бактерий, азотобактера, клубеньковых бактерий, у многих лучистых грибов, из которых готовят антибиотики. Фаги широко распространены в природе. Они находятся там, где встречаются бактерии и актиномицеты. Их выделяют из молока, почвы, воды, содержащих большое количество микробов. Основным источником фагов болезнетворных микробов являются больные люди и животные, а также бациллоносители. Фаги выделяются при кишечных инфекциях из испражнений, при гнойных заболеваниях из гноя. Особенно легко они выделяются в период выздоровления.
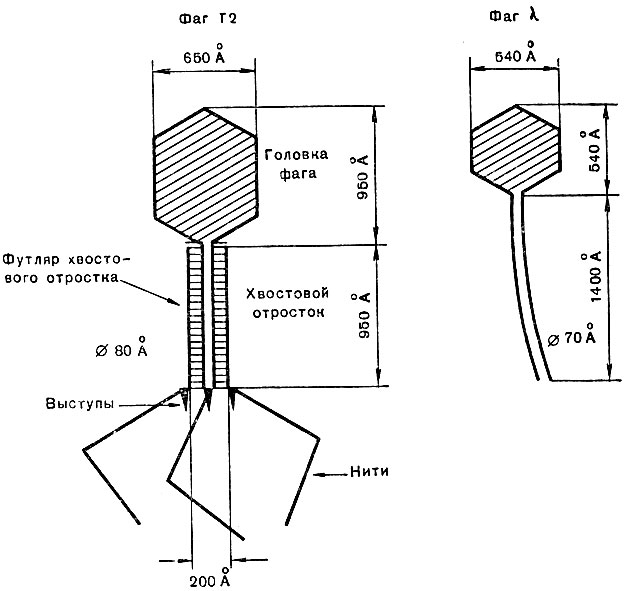
Рис. 47. Размеры и строение фага Т2
Для получения фага материал, содержащий его (вода, почва, испражнения, гной и пр.), высевают на жидкую питательную среду. В термостате на питательной среде вырастают бактерии, и в них начинают размножаться соответствующие фаги. Выросшую культуру фильтруют через бактериальный фильтр. Бактерии остаются на фильтре, а фаг переходит в прозрачный фильтрат. Если этот фильтрат прибавить к свежей бульонной культуре соответствующих бактерий, то фаг разрушит тела бактерий и культура посветлеет. На сплошном налете бактерий на твердой питательной среде наблюдаются пустые прозрачные круглые пятна. В этих пятнах находятся размножившиеся в клетках бактерий и лизировавшие их частицы фага. Такие пятна называются бляшками или негативными колониями фага. Если с этих бляшек петлей посеять в культуру чувствительного микроба, то опять произойдет явление бактериофагии.
Фаги более устойчивы к действию физических и химических факторов, чем неспороносные бактерии. В запаянных пробирках фаги могут сохраняться годами. Большинство фагов инактивируется при 65-75°. Фаги очень чувствительны к действию кислот и устойчивы к действию антибиотиков.
Важным свойством фагов является их специфичность. Каждый вид фага специфичен к определенному виду микробов. Часто наблюдается даже белее узкая специфичность внутри вида - типовая. Типоспецифический фаг действует не на все культуры данного вида бактерий, а только на некоторые. Так, брюшнотифозная бактерия в отношении брюшнотифозного фага разделяется на 44 типа.
Но специфичность фага относительна. Фаг можно адаптировать (приспособить) к паразитированию на другом виде бактерий путем многократных пересевов с клетками одного вида бактерий. Например, брюшнотифозный фаг можно адаптировать к дизентерийной палочке с наследственной потерей свойства лизировать брюшнотифозную палочку. Некоторые клетки чувствительной к фагу культуры могут приобрести устойчивость к разрушающему действию фага с передачей этого свойства по наследству. Образование фагоустойчивых культур бактерий происходит часто в результате мутаций.
Фаги проходят те же четыре фазы развития, что и вирусы. Фаг прикрепляется к клетке своим отростком, а не головкой. На конце отростка имеются длинные (130 ммк), но очень тонкие (2 ммк) белковые нити, которые улавливают в среде бактерии. Фаг прикрепляется к клетке бактерии особыми присосками на конце отростка.
В отростке фага имеется фермент типа лизоцима и молекула богатого энергией аденозинтрифосфата. Фермент разрыхляет оболочку бактерийной клетки, кончик отростка сжимается благодаря энергии АТФ и, как микрошприц, впрыскивает нуклеиновую кислоту фага в клетку. Белковая же оболочка остается снаружи бактерийной клетки и дальнейшего участия в развитии фага не принимает.
В следующей фазе, в первой ее половине, нуклеиновая кислота, проникшая в бактерийную клетку, не обнаруживается. В это время она, по-видимому, включается в генетический аппарат клетки и перестраивает клеточные механизмы синтеза, направляя их на производство фаговой нуклеиновой кислоты и фагового белка. О ходе созревания фагов в этот период стало известно в результате изучения сверхтонких срезов бактерийных клеток, инфицированных фагом, так как цельные клетки оказались очень толстыми для просматривания внутренних структур, в том числе и фагов, в электронный микроскоп. В тонких ультрасрезах было найдено, что сначала в различных участках протоплазмы клетки идет образование нуклеиновой кислоты, белковой головки и отростков. Затем, так сказать, детали в конце фазы объединяются и образуются зрелые фаги.
Клетки бактерий, несколько увеличившиеся от сформировавшихся внутри их фагов, внезапно взрываются и выбрасывают в окружающую среду до нескольких сотен молодых фагов. Полный цикл от адсорбции до выхода потомства фага из клетки занимает у разных видов фагов неодинаковое время - от 13 минут до 2 часов.
Освободившиеся из бактерийной клетки молодые фаги сразу начинают внедряться в другие клетки бактерий. Такой процесс происходит при благоприятных условиях, например, в пробирке с чувствительной культурой бактерий, до тех пор пока не будут лизированы все микробные клетки бактерий, пока не наступит стационарная фаза развития культуры.
При помощи метода меченых атомов установлено, что частицы потомства фага составляются из материала родительского фага, бактерийной клетки и питательной среды, на которой росла бактерийная клетка. Так, в фаговых частицах потомства содержится 35% фосфора от родительского фага, азота - 80% из питательной среды и 20% из бактерийной клетки.
Не всегда последняя фаза кончается разрушением бактерийной клетки фагом. При взаимодействии фага и клетки можно наблюдать самые разнообразные изменения клетки бактерии, не приводящие ее к гибели. Происходит изменение морфологии, вирулентности, биохимических свойств, приобретение фагоустойчивости и др. Фаги легко изменяются под влиянием внешних условий. Могут изменяться их морфологические, антигенные свойства: форма бляшек на твердых средах, адаптирование к другим типам и видам бактерий. Фаг легко подвергается мутационной изменчивости.
Но существует еще другой, особый тип взаимоотношений фага с клеткой. Как патогенные микробы могут иногда вызывать не только различной тяжести заболевания, но и скрытое, латентное течение заболевания, так и фаги, которые не вызывают гибели клеток, долгое время существуют в них в латентном (скрытом) состоянии. Такие фаги называются умеренными или неинфекционными. Взаимодействие умеренного фага с микробной клеткой выражается в лизогенизации бактериальной культуры. В лизогенных культурах наблюдается своеобразный симбиоз бактериальной клетки с фагом, при котором возможно сохранение и бактериальной клетки, и фага. Фаг не препятствует обменным реакциям и размножению бактериальных клеток в культуре. При делении бактериальной клетки фаг также переходит в обе новые клетки. Все это происходит в многочисленных поколениях. Фаг в них теснейшим образом связан с клеткой, но находится в неактивном, неинфекционном состоянии. Такой фаг А. Львов назвал профагом. Он не обнаруживается даже с помощью электронного микроскопа.
Поскольку в бактериальную клетку проникает только в основном фаговая ДНК, то очевидно, что умеренный фаг находится в клетке в виде фаговой ДНК, которая связана с ядерным аппаратом клетки - хромосомой, являясь как бы ее субъединицей.
Внешне бактериальная культура - скрытый носитель профага - ничем не отличается от нормальной, не зараженной фагом культуры. Но если эту лизогенную культуру профильтровать и фильтрат ее добавить к нелизогенной чувствительной культуре, то последняя будет лизироваться. Оказалось, что в единичных клетках лизогенной культуры, в одной из тысячи или десятка тысяч клеток, все же происходит постоянное превращение профага в зрелый фаг. Поэтому не происходит заметного внешнего изменения лизогенной культуры в нормальных условиях.
Профаг имеет свою генетическую информацию, необходимую для синтеза полноценных частиц данного вида фага. Это свойство профага проявляется, как только бактерии попадают в неблагоприятные условия. Было найдено, что под влиянием так называемых индуцирующих факторов - ультрафиолетового или рентгеновского облучения или воздействия химических веществ - происходит массовое превращение профага в активный фаг и клетки лизируются таким же образом, как и от воздействия инфекционного фага. По-видимому, индуцирующие факторы нарушают связь между генетическим аппаратом бактерии и профагом, а также активируют профаг.
Лизогения широко распространена в природе. У стафилококков, брюшнотифозных бактерий и у многих других почти каждый штамм является лизогенным. Большинство свежевыделенных культур от животных, растений и из почвы являются уже лизогенными.
Латентные, скрытые, вирусные инфекции встречаются не только у бактерий, но и у растений, животных и человека. По-видимому, бессимптомные вирусные инфекции встречаются в природе чаще, чем явные. Все без исключения растения картофеля сорта Король Эдуард заражены латентным вирусом, который не вызывает никаких признаков заболевания этого сорта картофеля, но вызывает резко проявляющееся заболевание у других сортов. Полиэдроз - вирусное заболевание шелковичных червей - внешне может не проявляться в течение всей жизни червя, но при изменении внешней среды, температуры, питания и пр. легко проявляется с выделением зрелой формы вируса. Вирусом герпеса человек заражается еще в раннем детстве. Но он проявляет себя только после охлаждения или после перенесения заболеваний гриппом и пр. в виде всем известных пузырьковидных высыпаний около губ и крыльев носа, содержащих прозрачную жидкость с вирусом.
Достижения в области вирусологии обнаруживают много сходного между вирусами и особенно фагами, с одной стороны, и опухолеродными вирусами - с другой. До сих пор еще нет определенных данных о причинах возникновения рака. По химической теории рак вызывается так называемыми канцерогенными веществами - главным образом продуктами неполного сгорания горючих веществ (каменного угля, нефти, сланцев, дыма, табака и пр.). В последнее время все более широкое признание получает вирусная теория. Имеется более 30 опухолей человека и животных, которые вызываются вирусами (саркома кур, папилома кроликов и др.), и число таких опухолей все время увеличивается (полиома мышей, лейкозы птиц, мышей и др.). Опухолеродные вирусы не разрушают клетки, а превращают их в опухолевые клетки, характерной особенностью которых является безграничное размножение.
Для победы над раком надо раскрыть основной механизм превращения нормальной клетки в злокачественную опухолевую. Этот механизм связан с нарушением синтеза белка, а следовательно, с изменениями в строении и функции нуклеиновой кислоты.
Фаг лизогенных культур может оказывать большое влияние на биологию бактерий, определять их свойства. Так, найдено, что некоторые фаги, выделенные из токсигенных лизогенных дифтерийных культур, при внедрении в нетоксигенные дифтерийные палочки превращают их в токсигенные, т. е. вырабатывающие дифтерийный токсин, вызывающий дифтерию человека (лизогенная конверсия). И это новое свойство уже передается по наследству. Фаги некоторых жгутиковых микробов могут вызывать у неподвижных бактерий образование жгутиков. Это явление называется трансдукцией.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'