Глава 5. Физиология растений. (П. А. Генкель, Е. М. Сенченкова)
Из всего периода полуторавекового существования науки физиологии растений XX в.- время ее наиболее бурного и многостороннего развития. Если в центре внимания физиологов прошлого столетия находились в основном вопросы воздушного и минерального питания, водного режима и дыхания растений, то такие проблемы, как физиология растительной клетки, ферментативная деятельность, рост, раздражимость, развитие, устойчивость к неблагоприятным условиям внешней среды, еще только зарождались. Теперь они выросли в большие области знаний, а их успешная разработка способствовала развитию не только физиологии растений, но и смежных с нею более молодых наук и дисциплин - биохимии растений, агрохимии, экспериментальной морфологии и экологии растений, физиологической анатомии, физиологии микроорганизмов.
Для развития мировой физиологии растений первых десятилетий XX в. характерно усиление биохимического подхода к решению многих проблем, особенно дыхания и фотосинтеза. В нашей стране начиная с 30-х годов характер физиологических исследований несколько изменился: изучение внутренней организации физиологических процессов в значительной мере уступило место вопросам экологии и растениеводства, сближающим фитофизиологию с насущными запросами земледелия. В странах Западной Европы развитие физиологии растений шло преимущественно в биохимическом направлении. Однако в конце 50-х годов советские физиологи растений возродили традиции А. Н. Баха, В. И. Палладина и С. П. Костычева, обратившись к углубленному изучению биохимических механизмов физиологических процессов.
За последние два десятилетия фитофизиология заняла ведущее место в ботанике. Об этом свидетельствует хотя бы увеличение удельного веса сообщений по физиологической проблематике на VIII - XI Международных ботанических конгрессах в Париже (1954), Монреале (1959), Эдинбурге (1964) и в Торонто (1969). Физиологический подход глубоко проник в такие ботанические дисциплины, как анатомия, цитология, фитоценология, фитопатология, микология, альгология и ряд областей прикладной ботаники. Даже морфология и систематика растений, всегда служившие классическим примером описательных наук, теперь по характеру и уровню экспериментов почти не отличаются от физиологии растений.
Изменился и характер самих физиологических работ. Совершенствование приемов исследований позволило фитофизиологам, не ограничиваясь описанием внешних проявлений ряда биохимических процессов, их исходных и конечных продуктов, вскрыть механизм различных форм обмена веществ, лежащих в основе питания, роста, размножения и реагирования на условия существования.
В современных фитофизиологических экспериментах широко используются методы радиоактивных и стабильных изотопов, различных видов хроматографии и электрофореза, приемов очистки и разделения на ионообменных смолах, ультрафиолетовой и инфракрасной спектроскопии, масс-спектрометрии, ультрацентрифугирования, электронной микроскопии, микрокалориметрирования и т. д. Успешному изучению физиологических процессов способствовало также создание специальных помещений искусственного климата - фитотронов, исключающих возможность случайного воздействия нежелательных метеорологических факторов и позволяющих быстро и точно изучать реакции растений в легко воспроизводимых и контролируемых условиях. Первый фитотрон был построен в 1949 г. при Калифорнийском технологическом институте в г. Пасадена. За последующие полтора десятилетия вступило в строй еще несколько сооружений такого типа, в том числе в 1957 г. в Институте физиологии растений им. К. А. Тимирязева АН СССР в Москве, а в 1963 г. во Французском национальном исследовательском центре в Жиф-сюр-Иветт.

Фитотрон в Жиф-сюр-Иветт (Франция)
Прогресс физиологии растений связан и с успехами смежных наук - биохимии, биофизики, цитологии и др. Так, данные, полученные биохимиками в 40 - 50-х годах о ферментных системах таких сложных процессов, как дыхание, брожение, фотосинтез и азотистый обмен, а также о принципах передачи энергии при различных формах обмена веществ, открыли перед физиологами растений большие возможности в исследованиях этих процессов. В настоящее время биохимия столь тесно сблизилась с физиологией растений, что разграничить эти области знаний иногда не представляется возможным.
Продолжая расширять и углублять традиционные связи с физикой и химией, биохимией и физической химией, анатомией и цитологией, экологией и микробиологией, агрохимией и почвоведением, физиология растений в последнее десятилетие вступила в тесный контакт с математикой, биофизикой, геохимией, генетикой и исследованиями на молекулярном уровне, что уже привело к решению некоторых вопросов, связанных с изучением роста, дифференциации, развития растений и действием на них веществ регуляторного характера.
Однако, несмотря на проникновение в тончайшие структуры и механизмы физиологических процессов, протекающих в органах, клетках и органеллах, в поле зрения исследователей остается весь растительный организм в целом.
Наряду с углубленной теоретической разработкой ряда проблем на молекулярном уровне в центре внимания физиологов по-прежнему стоят задачи решения таких практически важных вопросов, как обеспечение условий наиболее полного использования растениями солнечной энергии, углекислого газа, воды и элементов питания в целях повышения урожая и качества сельскохозяйственной продукции. В практику растениеводства все шире внедряются новые виды удобрения, ряд соединений для улучшения обмена веществ, комплексоны для борьбы с хлорозом, гербициды, регуляторы и активаторы роста. Установление зависимости образования и накопления веществ в растениях от определенных факторов среды позволяет вполне сознательно регулировать накопление углеводов и жиров, белков и клейковины, таннидов или терпенов, алкалоидов или глюкозидов и получать высокий выход этих веществ с единицы площади.
Успешная разработка многих практически важных вопросов, а также создание форм, устойчивых к низким температурам, засухе, засоленности почв и прочим неблагоприятным факторам были бы невозможны без знания филогенетической истории растений и закономерностей преобразования их наследственных свойств.
Сравнительная и эволюционная фитофизиология
Одной из характерных черт развития современной физиологии растений является широкое проникновение эволюционного подхода в решение все большего числа физиологических и биохимических проблем. Это нашло выражение в изучении процессов жизнедеятельности растения как единого организма, сохраняющего свою целостность на всем протяжении онтогенеза, в трактовке последнего как одного из звеньев филогенетической истории. Фитофизиологи стали разрабатывать вопросы происхождения и эволюции фотосинтеза, дыхания, различных ферментных систем и пр. Расширению подобных работ способствовало зарождение нового направления биохимических исследований - эволюционной биохимии.
Одним из действенных путей развития эволюционных принципов в физиологии растений по-прежнему остается экспериментальное изучение изменчивости растений, где плодотворные результаты дает использование сравнительного метода. Фитофизиологические исследования, проведенные на клеточном и субклеточном уровнях, позволили обнаружить у растений ряд важных приспособительных свойств, отвечающих специфическим особенностям их строения и условиям существования. Одновременно получены убедительные факты, свидетельствующие о тесном единстве растений со всем органическим миром (приложимость к растениям закона полярности, раздражимость, наличие гетеротрофности в питании зеленых растений, общность энергетических превращений в митохондриях растений и животных и т. д.). Все это позволяет сделать вывод, что современная физиология растений в целом вступила в новый период своего развития, характеризующийся переходом на уровень сравнительной и эволюционной физиологии.

Михаил Семенович Цвет (1872 - 1919)
Изучение фотосинтеза и хлорофилла
Центральной проблемой физиологии растений, как и в XIX в., оставался фотосинтез. Интенсивное развитие этой области знаний привело уже в первые десятилетия XX в. к возникновению таких автономных направлений, как химия пигментов, структура и биохимия пластид и продуктов фотосинтеза, экология фотосинтеза, фотосинтез различных групп растений и микроорганизмов, светокультура растений.
Начало века ознаменовалось рядом замечательных исследований в этой области. Для изучения химии хлорофилла большое значение имело открытие М. С. Цветом (1901) трех форм этого пигмента, названных им соответственно хлорофиллинами α, β и γ. В 1903 г. Цвет создал метод адсорбционного хроматографического анализа, который и позволил ему получить в чистом виде эти формы хлорофилла, четыре формы ксантофилла и другие пигменты. С 30-х годов хроматографический анализ стал находить все более широкое применение.
Пользуясь спектроскопическим методом, немецкий биохимик Р. Вильштеттер подтвердил существование двух форм зеленого пигмента, которые назвал хлорофиллом а и b (аналогичны хлорофиллинам α и β Цвета). Он впервые установил точный химический состав этих форм хлорофилла, каротина и ксантофилла, а также изучил состав более 40 производных хлорофилла. Вильштеттер обнаружил, что в состав зеленого пигмента входит не железо, а магний. Эти открытия (Нобелевская премия, 1915) означали первый шаг к осуществлению синтеза хлорофилла. Открытия были подытожены (совместно с А. Штолем) в монографии "Исследования хлорофилла" (1913), выход которой был крупным научным событием.

Рихард Вильштеттер (1872 - 1942)
Для изучения состояния хлорофилла в растениях большое значение имела гипотеза о существовании в хлоропластах хлорофилл - белкового комплекса, высказанная первоначально Цветом и затем развитая В. Н. Любименко (1921, 1923). Любименко назвал этот комплекс "естественным" хлорофиллом, в отличие от хлорофилла в молекулярном растворе различных вытяжек. К 50-м годам было точно установлено, что не только хлорофилл, но и другие пигменты растений и бактерий находятся в естественном состоянии в составе сложных пигменто - белково - липоидных комплексов.
В 30 - 40-е годы Г. Фишер с сотрудниками исследовали химическую природу зеленых пигментов пурпурных серобактерий - бактериохлоро-филла и зеленых бактерий - бактериовиридина, а Г. Стрейн и В. М. Мэнниг (1942, 1943) обнаружили новые формы зеленого пигмента - хлорофилл c у жгутиковых, диатомовых и бурых водорослей и хлорофилл b у красных водорослей. Изучение оптических свойств хлорофиллов а и b и их роли в фотосинтезе, начатое в 40-е годы Р. Эмерсоном, позволило ему и другим исследователям получить в 50-х годах ряд фактов, свидетельствующих о существовании двух форм хлорофилла а, каждая из которых в свою очередь является составной частью двух пигментных систем. С 50-х годов развернулись исследования роли каротиноидов в фотосинтетических процессах.
Выявление основных этапов биосинтеза хлорофилла связано с исследованиями Т. Н. Годнева, С. Граника, Л. Богорада, Д. Смита, Д. Шемина и других, начатыми еще до 50-х годов. Общими усилиями удалось установить, что образованию как хлорофилла, так и гема, входящего в состав гемоглобина, предшествует синтез протопорфирина, после чего пути их биосинтеза расходятся: включение в молекулу протопорфирина атома железа дает гем, а включение магния приводит через ряд превращений к образованию хлорофилла.
Еще в 30-х годах Г. Фишер предпринял попытку синтезировать хлорофилл вне растения, но большего успеха добились в 1960 г. его ученики М. Штрель, А Колоянов и Г. Коллер, получившие искусственно феофорбид - основу молекулы хлорофилла без магния и фитола. Одновременно такой же синтез осуществил американский химик Р. Вудворд (1960; Нобелевская премия, 1962), который получил хлорофилл.
Кинетика и химизм фотосинтеза
С начала XX в. существенно изменился характер исследований зависимости фотосинтеза от внешних условий. Влияние на фотосинтез количества и качества света, концентрации углекислого газа и кислорода воздуха, влажности, температуры и других факторов стало изучаться в комплексе, а не изолированно. Стали учитывать также внутренние факторы - концентрацию хлорофилла, накопление ассимилятов, возраст листьев и др. Мысль об одновременном учете основных сопряженных факторов получила развернутое экспериментальное и теоретическое обоснование в учении английского фитофизиолога Ф. Блэкмана (1905) о лимитирующих факторах фотосинтеза, уточненном позднее Р. Хардером (1921) и Г. Лундегордом (1921, 1924, 1937).
Многочисленные исследования, посвященные анализу воздействия внешних и внутренних факторов на воздушное питание растений, породили новое направление - экологию фотосинтеза, основным содержанием которого стало изучение этого процесса в различных природных условиях. Это направление получило особенно широкую разработку в трудах советских физиологов.
В 1905 г. Ф. Блэкман и Г. Маттеи установили, что процесс фотосинтеза складывается из двух фаз: первая включает световые (фотохимические) реакции, не зависящие от температуры, вторая - темновые (химические) реакции, тесно связанные с температурным фактором. Обе фазы фотосинтеза тщательно исследовал немецкий биохимик О. Варбург (1919, 1920). Он показал, что темновые (он предложил называть их блэкмановскими) реакции фотосинтеза можно изменить действием различных температур. Применив прерывистое освещение, Варбург определил продолжительность обеих фаз.
Многочисленные исследования были направлены на выяснение характера превращений веществ и энергии на обоих этапах фотосинтеза. Опираясь на открытие X. Виландом (1913) процессов окисления за счет дегидрогенизации воды (см. главу 6), Т. Тунберг (1923) и Ф. Вейгерт (1923, 1924) высказали мысль, что первоначальный, фотохимический акт фотосинтеза заключается в разложении воды на водород и перекись водорода, а на последующем нефотохимическом этапе водород восстанавливает углекислый газ до углеводного соединения. Более обстоятельно гипотеза окисления воды в ходе фотохимического этапа фотосинтеза была развита К. Б. Ван-Нилем (1931-1935) и Г. Гаффроном (1939 - 1944), открывшим явление фоторедукции.
Новые перспективы фотохимических исследований фотосинтеза открылись после опытов английского биохимика Р. Хилла (1937, 1939), в которых изолированные хлоропласты растертых листьев эффективно восстанавливали на свету хиноны и выделяли кислород. При этом в качестве водородного акцептора был использован не СО2, а железо-калиевый оксалат. Эта реакция, получившая название реакции Хилла, подтвердила выводы Ван-Ниля и Гаффрона о том, что процессы восстановления СО2 и выделения О2 независимы и их можно изучать каждый в отдельности. Кроме того, она опровергла мнение, будто изолированные выделенные из клетки хлоропласты не способны осуществлять фотосинтез. Позднее было установлено, что фотосинтезирующая способность зеленых пластид сохраняется в том случае, если их поместить в водную среду, содержащую всю совокупность ферментов зеленой клетки.
Метод меченых атомов позволил одновременно советским (А. П. Виноградов, Р. В. Тейс, 1941) и американским (С. Рубен с сотрудниками, 1941) исследователям окончательно доказать, что выделяемый при фотосинтезе растений кислород образуется не за счет С02 или других окислителей, как это имело место в реакции Хилла, а за счет фотолиза воды.
Для изучения световой фазы фотосинтеза многое сделали в 40-х годах. А. Н. Теренин, а также Г. Н. Льюис и М. Каша, исследования которых разъяснили вопрос о первичной фотореакции хлорофилла и трансформировании им световой энергии*.
* (См.: А. Н. Теренин. Фотохимия хлорофилла и фотосинтез. М., Изд-во АН СССР, 1951.)
Новые методы исследования позволили выявить в процессе фотосинтеза три основные стадии: 1) отщепление атомов водорода под действием света от молекулы воды с выделением молекул кислорода; 2) перенос атомов водорода в ходе двух фотохимических реакций и 3) использование тех же атомов водорода для превращения углекислоты в сахара (темновые реакции).
Из трех названных стадий фотосинтеза наименее изучена первая. Несколько полнее исследована вторая, на которой благодаря хлорофиллу, играющему роль фотокатализатора, происходит аккумуляция световой энергии. За этим основным фотохимическим процессом следует ряд вторичных, так называемых темновых реакций, в которых накопления энергии не происходит. Суть же совершающихся при этом окислительно-восстановительных реакций заключается в переносе атомов водорода или электрона, образующихся на первой стадии фотосинтеза, от одной молекулы к другой.
Представление о том, что вторая стадия фотосинтеза состоит из двух фотохимических реакций, связанных с возбуждением упомянутых выше двух пигментных систем, к настоящему времени получило серьезное экспериментальное обоснование и стало почти общепризнанным. Однако механизм этих реакций раскрыт пока еще неполностью. Известно, что энергия, накопленная второй системой, используется для образования аденозинтрифосфата (АТФ), а первая система за счет энергии света обеспечивает восстановительные синтезы водородом. В итоге взаимодействие двух фотохимических систем приводит к образованию АТФ, выделению кислорода и синтезу органических веществ из углекислого газа. Подробнее с механизмом происходящих при этом процессов, нашедщих свое выражение в ряде схем, можно ознакомиться в ряде работ 60-х годов Д. Арнона, Е. Рабиновича, В. П. Скулачева и др.
Третья стадия фотосинтеза, или так называемый цикл углерода, изучена наиболее полно. Крупнейших успехов в этой области достигли американский биохимик М. Кальвин с сотрудником А. Бенсоном и др. В начале 50-х годов было показано, что первым стойким продуктом фотосинтеза является фосфоглицериновая кислота, а акцептором углекислого газа - рибулезодифосфат. В итоге Кальвин (Нобелевская премия, 1961) разработал схему превращения углерода в процессе фотосинтеза, основное содержание которой сводится к следующему. Рибулезодифосфат, присоединяя молекулу СО2, распадается на две молекулы фосфоглицериновой кислоты, а последняя подвергается восстановлению при участии АТФ и пиридиннуклеотидов. Дальнейшие сложные превращения приводят к образованию конечных продуктов (углеводов) и регенерации рибулезодифосфата, Эту стадию фотосинтеза называют также циклом Кальвина. В настоящее время изучены не только вещества, образующиеся на отдельных этапах данной стадии, но и ферментные системы, катализирующие различные реакции цикла.
Процесс образования АТФ при фотосинтезе в отличие от синтеза АТФ при дыхании (окислительное фосфорилирование) Л. Арнон (1954) назвал фотофосфорилированием. Было установлено, что использование АТФ в качестве "единой энергетической валюты" идет в основном по разным руслам. Образующийся при фотофосфорилировании АТФ используется в первую очередь в реакциях восстановления углекислого газа и только частично для вторичных синтезов. АТФ же, синтезируемый при дыхании, используется преимущественно для других процессов жизнедеятельности растений - поддержания физико-химического состояния клеточных структур, осмотических явлений, движения протоплазмы, синтеза различных сложных соединений и др. Примечательно, что источник для синтеза АТФ и при фотосинтезе, и при дыхании как в растительной, так и в животной клетке один и тот же (см. главу 6).
Д. Арнон с сотрудниками, открывшие в 1954 г. процесс фотосинтетического фосфорилирования на изолированных хлоропластах, многое сделали для выяснения условий, необходимых для обеспечения более высокой эффективности преобразования световой энергии в энергию химических связей АТФ, и выявления особенностей этого процесса.
Управление фотосинтезом
В 50-х годах А. А. Ничипорович (1955, 1959) совместно с сотрудниками разработали представление о разнокачественности или разнообразии основных продуктов фотосинтеза и о возможности образования на свету как углеводов, так и белков с преобладанием синтеза тех или иных продуктов в зависимости от внешних (снабжение азотом, количество и качество света) и внутренних (возраст растений, наследственные свойства и т. д.) условий. Знание этих условий дает возможность управлять образованием этих продуктов при фотосинтезе.
Большую роль в разработке вопроса об управлении деятельностью фотосинтетического аппарата играют современные исследования явления адаптации фотосинтеза растений применительно к различным условиям и особенно количеству и качеству света. Они показали, что адаптивная перестройка фотосинтетического аппарата может совершаться относительно быстро и сопровождаться изменениями как в структуре и биохимических особенностях этого аппарата, так и в изменении активности и качественной направленности его работы. Довольно широко изучена адаптация фотосинтеза к таким факторам среды, как интенсивность, спектральный состав света, уровень снабжения элементами минерального питания, углекислым газом и кислородом, а также сопряженность процесса фотосинтеза с другими физиологическими процессами.
Проблема фотосинтетической продуктивности наиболее активно стала разрабатываться с конца 40-х годов в Советском Союзе под руководством А. А. Ничипоровича, а также в Японии, Англии, США, Нидерландах и других странах. На основе этих исследований значительно расширилось и приобрело полную самостоятельность учение о сложных фотосинтезирующих системах (включая условия и факторы высокой продуктивности). Новая проблема включает в себя определения фотосинтетически активной радиации в различных географических зонах и степени полноценности этой радиации для фотосинтеза в зависимости от температуры, влажности, почвенного плодородия, физиологических и экологических особенностей растений. Предметом изучения стали также закономерности структурной и пространственной организации таких фотосинтетических систем, как естественные растительные сообщества и различные посевы. Исследуются их оптические свойства и создаваемые в них световые режимы, определяются несоответствия между теоретически возможными и реальными показателями фотосинтетической деятельности, выясняются причины этих несоответствий и пути их преодоления. Все это подчинено одной цели - найти условия для повышения фотосинтетической активности растений в посевах на единицу площади с 1 - 3 до 5 - 6% от падающей солнечной энергии, что привело бы к увеличению накопления органической массы и повышению урожайности.
Разработка теории фотосинтетической продуктивности растений основывается также на культивировании растений в закрытом грунте при искусственном свете. Широкое развитие светокультура получила в СССР, в США и Франции. Использование ксеноновых ламп позволило в два раза повысить урожай и вдвое ускорить сроки созревания в сравнении с лучшими условиями открытого грунта. Коэффициент использования света этих ламп растениями достигает 8%. Качественный состав света при выращивании в современных теплицах базируется на результатах исследования зависимости фотосинтеза от спектрального состава света.
Широкое развитие в 60-х годах получили работы по массовой культуре одноклеточной зеленой водоросли - хлореллы. В специальных, автоматически действующих аппаратах, куда помещали культуры хлореллы, исследователям разных стран удалось получить значительно более высокий коэффициент использования световой энергии, чем это пока достигнуто в полевых или тепличных условиях для высших растений, и различное содержание белков, жиров и углеводов в биомассе разных штаммов.
Одновременно развернулась успешная работа по использованию фотосинтеза растений в точно контролируемых и автоматически управляемых замкнутых системах. Так, В. Е. Семененко с сотрудниками в сконструированной ими установке по непрерывному выращиванию хлореллы получили фотосинтетической продукции в 25 - 30 г сухого вещества на 1 л суспензии в сутки при КПД лучистой энергии 6 - 8%. Подобные фотосинтезирующие системы, способные поглощать из воздуха большие объемы углекислого газа и выделять столь же много кислорода, в будущем, возможно, окажутся спутниками человека в длительных космических полетах (см. главу 28).
Природа дыхания
К началу XX в. о дыхании сложилось представление как о процессе, противоположном фотосинтезу, однако о его внутреннем механизме почти ничего не знали. Существенный вклад в разрешение этой проблемы внесли русские ученые, начиная с А. Н. Баха и С. П. Костычева.
Благодаря усилиям исследователей разных стран (В. И. Палладин, О. Варбург, Д. Кейлин, Г. Виланд, А. Сцент-Дьёрдьи, Г. А. Кребс и др.) в 50-х годах сложились современные представления о биохимической природе процесса дыхания. Было доказано, что в основе дыхания как растительных, так и животных тканей лежат по существу одни и те же окислительные реакции и энергетические процессы (см. об этом в главе 6).
Согласно современным представлениям, дыхание - это сложная система сопряженных окислительно-восстановительных процессов, в ходе которых происходит химическое преобразование углеводов, белков и жиров и использование высвобождающейся при этом энергии.
В 1957 г. Г. Л. Корнберг и Г. А. Кребс открыли новый дыхательный цикл глиоксалевой кислоты. Он представляет модификацию цикла лимонной кислоты и лежит преимущественно в основе дыхания бактерий и грибов, осуществляясь за счет превращения жировых веществ в углеводы.
В 50-х годах было начато изучение у растений еще одного цикла окисления углеводов, апотомического, или гексозомонофосфатного, открытого в 30-х годах у дрожжей и в тканях животных. Сравнение апотомического дыхательного цикла с фотосинтетическим циклом восстановления СО2, предложенным М. Кальвином (1957), показало, что первый представляет собой полное зеркальное повторение второго, т. е. протекает в обратную сторону. Открытие этого факта дает основание предполагать о существовании тесной связи этих двух процессов в организме растений*.
* (См.: О. А. Семихатова. Смена дыхательных систем. Критический анализ методов исследования. Л., "Наука", 1969.)
Большая серия исследований была посвящена изучению зависимости дыхания от особенностей растительных организмов, от других протекающих в них процессов жизнедеятельности и различных факторов внешней среды. В. Стайльз и В. Лич в 50-х годах установили, что растениям различных систематических групп свойственна определенная интенсивность дыхания, а А. Сцент-Дьёрдьи еще в 30-х годах обратил внимание на то, что различным растениям присущ даже определенный характер окислительной системы. Так, однодольные, а из двудольных крестоцветные и тыквенные, полностью или частично лишены полифенолоксидазной активности. Теперь считается вполне достоверным, что особенности дыхательной системы различных растений отражают их положение в филогенетической системе. Накапливаются также данные о различиях в дыхательной активности у различных органов растений, о зависимости дыхания от возраста. Имеются основания предполагать, что в ходе онтогенеза процесс дыхания может изменяться не только количественно, но и качественно.
Получены интересные данные о связи дыхания с фотосинтезом, минеральным и водным питанием, биосинтетической функцией клеток и ростом растения. В течение многих десятков лет было принято противопоставлять дыхание фотосинтезу: их рассматривали как два прямо противоположных по своему биологическому значению процесса - ассимиляцию и диссимиляцию. Теперь, когда между ними установлена тесная связь, фотосинтез и дыхание могут служить яркой иллюстрацией кардинального положения материалистической диалектики о единстве противоположностей. Однако характер связи между дыханием и фотосинтезом выяснен еще не полностью. Это относится в первую очередь к взаимосвязи ферментативных механизмов обоих процессов.
Изучению зависимости поглощения ионов корнями растений от аэробного дыхания тканей посвящена серия работ шведского исследователя Г. Люндегорда, сформулировавшего анионную теорию дыхания (1955), которая до сих пор дискутируется. Начаты исследования механизма использования энергии АТФ в процессах солевого питания, а также вопроса о природе первичных акцепторов поглощаемых клеткой ионов.
Современные представления о взаимосвязи дыхания и потребления растением воды подробно освещены в работе Д. П. Хаккетта (1959). Автор разделяет точку зрения об активности процесса поглощения воды растительными тканями, который через посредство ауксинов сопряжен с дыханием, поставляющим для него энергию. Согласно данным Г. Бурстрема (1953), поглощение воды связано с процессами дыхания не менее сложной зависимостью, нежели поглощение минеральных солей.
Минеральное питание
Проблема минерального питания растений в XX в. почти целиком стала достоянием агрохимии, составив основу учения об удобрениях. Существенный вклад в это учение внесли отечественные физиологи растений и прежде всего Д. Н. Прянишников (Ленинская премия, 1926) и его школа. Ниже будут затронуты лишь специальные, физиологические вопросы минерального питания растений.
Успешное развитие проблемы стало возможным благодаря совершенствованию методов исследования и прежде всего техники и организации вегетационных опытов. Для постановки таких опытов еще в конце прошлого века стали сооружать не только отдельные вегетационные домики, но и целые павильоны.
В 1900 г. французский агроном П. Мазэ начал выращивать растения в стерильных условиях, удаляя микроорганизмы, способные изменять состав питательной среды. Методика таких "стерильных культур" в дальнейшем получила широкое распространение. Кроме того, для сохранения постоянства состава питательных растворов на всем протяжении культивирования стали практиковать так называемые "текучие растворы" (П. С. Коссович, 1902). Изучение ионной концентрации питательных растворов позволило создать и поддерживать постоянным их оптимальный состав, что имело, в частности, важное теоретическое и практическое значение для производственного выращивания растений в теплицах гидропонным способом. Вегетационные опыты проводились в водных и песчаных или гравийных культурах.
К началу XX в. было твердо установлено, что питательные растворы должны включать в себя семь необходимых элементов: азот, фосфор, серу, калий, кальций, магний и железо (углерод, водород и кислород растение получает в основном из воздуха и воды). Однако для создания научных основ учения о минеральном удобрении нужно было знать, в каких соединениях растения способны усваивать эти элементы. В отношении азотного питания растений эта задача была решена в работах Д. Н. Прянишникова, итог которых подведен в книге "Азот в жизни растений и в земледелии СССР" (1945).
С изучением азотного питания растений были тесно связаны исследования способности клубеньковых бактерий фиксировать азот воздуха (М. В. Бейеринк, 1901; Л. Гильтнер, К. Штермер, 1903; Ф. Ленис, Р. Ганзен, 1921; М. Экхард с сотрудниками, 1931; и др.). Они позволили установить существование на корнях бобовых растений около 20 рас клубеньковых бактерий. Было обнаружено, что эффект азотфиксации зависит от подбора симбиотических пар: с одной стороны, бобового растения, а с другой - того или иного штамма бактерий. На этой основе еще до 30-х годов был выполнен большой объем работ по изучению применения культуры азотобактера для повышения урожая различных культурных растений. Обнаружение клубеньков на корнях ольхи и лоха позволило предположить, что симбиоз такого рода - явление, в той или иной мере присущее всему растительному миру.
С рассматриваемым периодом связана постановка и частичное решение вопроса о роли микроэлементов в жизни растений. До 20-х годов опыты в данном направлении носили случайный характер, но уже и в них было обнаружено стимулирующее действие на рост растений марганца, цинка, кобальта, никеля, бора, мышьяка и меди. Считавшиеся до того токсичными, эти элементы в малых дозах улучшали развитие растений, почему в дальнейшем они стали использоваться в качестве добавок к удобрениям. В 30 - 40-е годы была окончательно доказана необходимость для растений марганца, цинка, меди, бора и молибдена. Их выделили в группу основных микроэлементов. В особую группу были объединены галлий, кремний и алюминий, необходимость которых доказана пока лишь в отношении некоторых растений. К 50-м годам в составе растений было обнаружено около 40 микро- и ультрамикроэлементов. Но их необходимость для жизни растений была установлена не для всех микроэлементов.
С физиологической точки зрения все необходимые для растений элементы, получаемые из почвы, были условно разделены на две группы. В первую группу вошли металлоиды (азот, сера, фосфор, кремний, хлор, иод и др.), поступающие в растение в виде анионов солей, в другую - металлы (калий, кальций, магний, железо и др.), поступающие в виде катионов. С несомненностью установлена специфичность катионов металлов, т. е. невозможность замены одного другим, но полной ясности в представлениях об их функциях еще нет.
Предметом пристального внимания исследователей стала в XX в. проблема внекорневого питания растений, название которой предложили в 1903 г. русские энтомологи И. Я. Шевырев и С. А. Мокржецкий. В целях лечения и борьбы с вредными насекомыми они вводили различные вещества под кору деревьев и считали возможным таким же способом вводить и питательные вещества. Позднее понятие о внекорневом питании было расширено. Под этим понятием в практику вошли не столько инъекции в ствол, сколько опрыскивание растворами минеральных веществ листьев и других наземных органов как древесных, так и травянистых растений. Первые опыты в этом направлении провел немецкий агрохимик Л. Хильтнер (1909, 1912). Он опрыскивал картофель растворами солей азота, калия, магния и даже гумуса. В 30-х годах исследования по внекорневому питанию растений развернулись также в лаборатории Д. Н. Прянишникова, а позднее и в полевых условиях. Большая литература, накопившаяся к настоящему времени по вопросу о внекорневом питании, свидетельствует о его высокой эффективности как средства подкормки. Оно особенно оправдало себя на культурах свеклы и хлопчатника.
Поступление веществ в растение
Исследования процесса усвоения минеральных веществ корнями всегда стояли в тесной связи с изучением механизма поступления, передвижения и превращения этих веществ в ходе обмена. При этом прежде всего учитывали характер среды, на которой произрастает растение. Разработка метода определения концентрации водородных ионов (рН) в начале 20-х годов позволила установить зависимость корневого питания растений от рН почвенных растворов (С. Аррениус*, 1922; Д. Н. Прянишников, 1931).
* (За разработку теории электролитической диссоциации С. Аррениус был удостоен в 1903 г. Нобелевской премии.)

Дмитрий Николаевич Прянишников (1865 - 1948)
В тесной связи с изучением поступления питательных веществ в корни и оттуда в другие части растений, а также выделения продуктов внутриклеточного метаболизма находились исследования клеточной проницаемости (см. об этом в главе 10).
Уже в начале XX в. существовало мнение (Е. Овертон, А. Натансон), что поглощение веществ живыми клетками должно быть тесно связано с другими процессами жизнедеятельности растений, но экспериментальное обоснование эта идея получила лишь спустя четверть века. Одним из первых исследователей, показавших существование взаимообусловленности в деятельности наземных органов и корня, наличие всесторонней и глубокой связи минерального питания с процессами обмена веществ, структурой плазмы, ростом, формообразованием и развитием растений был Д. А. Сабинин (1925). Он установил, что первым этапом поглощения минеральных солей является их адсорбция на поверхности корневой системы - положение, получившее затем всеобщее признание. В 1929 г. он выдвинул положение о том, что корневая система растений является органом не только поглощения, но и превращения веществ. В 40-х годах И. И. Колосов выявил значение обменной адсорбции для поглощения корнями как катионов, так и анионов. Были получены также интересные данные о зависимости поглощения питательных веществ от интенсивности дыхания.
Чешский ботаник С. Прат (1923) показал, что между количеством испаренной растением воды и количеством солей, поглощенных корнями из почвенного раствора, прямой связи не существует. В 20-х годах было установлено также, что ионы могут передвигаться по растению независимо от транспирационного тока воды по всем направлениям и даже навстречу этому току. О. Ф. Туева (1926) показала, что ионы солей, передвигаясь в направлении, противоположном движению воды в корневой системе, могут выделяться из растения наружу. В 30 - 40-х годах Г. Иенни, Р. Оверстрит, А. В. Петербургский и другие ученые разработали теоретические представления о механизме так называемого контактного обмена ионов между корневыми волосками и коллоидными частицами твердой фазы почвы. Однако в последующие годы появились соображения, что абсолютизировать независимость поглощения минеральных солей от почвенной воды и транспирации не следует.
К началу 50-х годов господствующее положение получила ультрафильтрационная теория поступления веществ в клетку. Другую, сорбционную теорию активно поддерживал А. С. Трошин (1953, 1956). При участии и помощи Д. Н. Насонова он собрал и проанализировал большой материал, свидетельствовавший о несостоятельности ультрафильтрационной теории. В результате в 50-е годы разногласия в трактовке механизма поступления веществ в клетку не сгладились, а углубились. Однако, несмотря на бесспорность ряда положений сорбционной теории, заметного влияния на развитие представлений о поступлении питательных веществ в живую клетку она не оказала.
В последние десятилетия стало очевидным, что в основе как поверхностной, так и внутренней структуры цитоплазмы и ее органелл лежит мембранный принцип. Электронно-микроскопические исследования позволили установить трехслойность строения элементарной мембраны. В результате в новом свете возродилась мембранная теория поглощения.
Для современных исследований поглощения минеральных веществ корнями растений характерно вычленение двух качественно различных процессов этого явления - метаболического, или активного, и неметаболического, или пассивного. Для первого характерно включение в обмен веществ элементов уже в момент их поглощения. Движущей силой второго является диффузия и осмос. Об активности поглощения ряда ионов свидетельствуют многочисленные исследования, выполненные в 40 - 50-е годы. Вместе с тем некоторые авторы (А. Уоллес, 1966) приводят достаточно убедительные данные о неметаболическом поглощении некоторых катионов.
Объяснить механизм неметаболического способа поглощения веществ помимо диффузии, не играющей существенной роли в поглощении веществ живой плазмой, позволяет пиноцитоз - прерывистое захватывание капелек жидкости из окружающего раствора, открытое В. Льюисом (1931) у животных клеток, и Р. Бюва, Г. Паладом и У. Джонсоном у клеток растений. Вместе с тем становится очевидным, что неметаболическое поглощение веществ - это не пассивный, а активный физиологический процесс, обусловленный интенсивностью движения протоплазмы и степенью деформации ее поверхности. В 50-е годы идея о зависимости поглощения и передвижения веществ от метаболизма в целом получила развитие в работах А. Л. Курсанова. За последние 10 - 15 лет основное внимание исследователей было обращено на изучение метаболического пути поглощения минеральных веществ и связь этого процесса с дыханием растений.
Для объяснения механизма активного поглощения минеральных элементов, непосредственно связанного с обменом веществ поглощающих клеток, в последнее время предложен ряд гипотез. Наибольшим признанием пользуется так называемая теория переносчиков, впервые сформулированная В. Остергаутом и У. Стенли (1932). В настоящее время многие ученые широко используют эту гипотезу для объяснения механизма поглощения и транспортировки не только катионов, но и анионов, и органических веществ.
Большой экспериментальный материал накоплен по вопросу о зависимости поглощения питательных веществ корневой системой растений от внешних условий. Глубокий интерес к выяснению зависимости поглощения питательных веществ от их природы, соотношения отдельных компонентов, влажности, аэрации, температуры и кислотности почвы обусловлен главным образом потребностями практики. Такие исследования необходимы для разработки и рационализации агротехнических приемов и создания эффективной системы удобрений.
В последние годы много внимания уделяется изучению роли микроэлементов в жизни растений. Интерес к этому вопросу связан не только с его большой значимостью для сельскохозяйственной практики, но и важностью для теоретической биологии. Обнаружение одних и тех же микроэлементов (Mg, Zn, Cu, Fe, Co и др.) у растений и животных свидетельствует об их единстве. В немалой степени этому способствовало обнаружение микроэлементов в составе большого числа ферментов обоих царств органического мира.
В 50 - 60-е годы учение о металлоферментах стало одной из центральных проблем биохимии и физиологии. Многочисленные исследования касаются механизма усиления микроэлементами каталитического действия присоединения металлов с протеином. Интересные данные получены также о связи микроэлементов со стимуляторами роста и витаминами.
Совсем недавно было выяснено, что в превращении материи и энергии в растительном организме принимают непосредственное участие не отдельные элементы, а все минеральные вещества. Их роль с несомненностью установлена в процессах фотосинтеза, дыхания, водообмена, роста и развития. Эти вещества являются или структурными компонентами функциональных систем (фотосинтетического аппарата, дыхательной цепи, центров синтеза белка и других важных соединений) или составной частью ферментных систем, ответственных за осуществление физиологических процессов, или, наконец, их кофакторами. Таким образом, с 50-х годов все более укрепляется убеждение в том, что фотосинтез, дыхание, рост, развитие и минеральное питание - это нераздельные и взаимообусловленные процессы образования и превращения живого вещества растительного организма.
Передвижение веществ и их обмен
Для объяснения причин передвижения питательных веществ по растению в XX в. был выдвинут ряд теорий, из которых упомянем следующие. Немецкий физиолог Е. Мюнх (1930) утверждал, что движение органических веществ в проводящей системе флоэмы определяется наличием градиента тургорного давления, а американский исследователь О. Картис (1937) связывал это явление с круговым движением протоплазмы в клетках ситовидных трубок. Однако опыты не подтвердили этих теорий. Не выдержали экспериментальной проверки и представления о диффузной природе передвижения веществ. Недостаток всех подобных теорий заключался в том, что в их основу были положены чисто физические причины.
С иных позиций подошел к объяснению передвижения веществ Д. А. Сабинин (1949). Он видел причины существования в растении одностороннего тока воды и растворимых веществ в физиологической полярности клеток, обусловленной различиями в обмене веществ между разными участками протоплазмы.
В начале XX в. развернулись исследования цикличности превращений различных элементов в обмене веществ растений. Наибольшие успехи были достигнуты в раскрытии картины азотного и фосфорного обмена (Д. Н. Прянишников). Был исследован механизм фиксации аммиака и его участие в органических соединениях растений. Применение меченых атомов азота позволило Г. Хевеси и его сотрудникам (1940) показать, что между различными органами и тканями растения существует непрерывный обмен азотистых соединений, а последующие эксперименты обнаружили постоянное обновление тканевых белков.
Исследованиям фосфорного обмена в растениях положил начало Л. А. Иванов (1901, 1906). Ему удалось проследить пути превращения фосфора в прорастающих и созревающих семенах. Особое внимание он уделил нуклеопротеидам, образование которых, как он отмечал, всецело зависело от фосфорной кислоты. Им же были установлены факты, положенные затем в основу современных представлений о фосфорилировании Сахаров в процессах дыхания и брожения. В результате дальнейших исследований выяснены некоторые детали фосфорного обмена в растениях, в частности, тот факт, что в отличие от азота и серы соединения фосфора всегда, при всех их изменениях в растениях остаются окисленными. Ряд исследований, особенно на созревающих семенах, показал, что ассимиляция серы растением выражается в восстановлении поглощенных сульфатов и синтезе аминокислот и белков. Характер превращений других элементов и их место в обмене веществ у растений пока еще полностью не раскрыты.
Для изучения обменных процессов и их регулирования в клетке существенное значение имела разработка в 40-х годах Ф. Уайтом, П. Нобекуром и Р. Готре метода культуры изолированных органов и тканей на жидкой или твердой питательной среде.
В настоящее время разработаны не только методы культуры изолированных тканей, но и метод культуры суспензий клеток в жидкой питательной среде. Применяя метод культуры изолированных клеток тканей моркови, Ф. Стьюард (1958), а затем Р. Г. Бутенко (1964) получили регенерацию целого растения.
Водный режим
Прогресс физиологии растений в XX в. коснулся и такой важной в практическом отношении проблемы, как водный режим, включающей в себя вопросы поглощения воды, перемещения ее по растению и испарения в окружающую среду, а также выявление главных типов растений по их отношению к воде.
Изучение механизма поступления воды в растение велось на основе исследований осмотических свойств растительных клеток, в частности, клеток корневой системы. Опыты А. И. Броуна (1907, 1909), Г. Коллинса (1918), А. А. Гурвича (1929) и других выявили полупроницаемость клеточных оболочек, довольно легко пропускающих воду, но препятствующих проникновению растворенных в ней веществ. Оказалось, что данное свойство оболочек сохраняется даже после убивания протоплазмы. До 20-х годов была также установлена полупроницаемость покровов семян различных групп растений и доказано, что оболочки растительных клеток, как правило, не оказывают сопротивления диффузии воды.
Немецкие исследователи А. Уршпрунг и Г. Блюм (1920) развили представление о сосущей силе - способности клеток поглощать воду из окружающей среды благодаря разности концентраций растворов внутри и вне их. Ими же были разработаны первые методы ее определения. В 1936 г. Т. А. Беннет-Кларк с сотрудниками установили явление активной секреции воды из протоплазмы в вакуоли клеток под действием электроосмотических сил, но лишь в 1946 г. они смогли разработать способ определения величины электроосмотического компонента внутриклеточного давления.
Хотя ни у кого не вызывало сомнений то, что основными органами, доставляющими растению воду, являются корни, первые обстоятельные исследования корневой системы были проведены лишь в 10-х годах. Тогда были получены ценные данные о скорости роста, глубине проникновения и степени развития корневых систем, а также скорости продвижения по ним воды у различных растений. Определения Г. Дитмера (1937) убедительно показали, что общая поверхность корневой системы растения, как правило, в десятки раз превышает поверхность наземных органов. Изучение зависимости поглощения воды корнями от внешних условий, начатое еще в прошлом веке, в XX столетии развернулось очень широко, охватив действие температуры, аэрации, кислотности почв и других факторов.
Несмотря на двухвековую историю изучения передвижения воды по растению, вопрос о причинах этого явления не был решен, и в начале XX в., вплоть до 30-х годов, велся спор о том, принимают ли участие в передвижении воды живые клетки растений. В анализе причин, вызывающих поднятие воды от корней до вершины дерева, в частности Г. Г. Диксон, Е. Овертон и Г. Молиш (20-е годы), опирались на непрерывность водных нитей в сосудах и наличие сил сцепления, отрицая роль живых клеток. При этом движущие силы они усматривали в различии величин давления диффузии воды между почвой, растением и окружающей атмосферой.
Для изучения водного тока в ксилеме были использованы методы инъекции красок, замораживания тканей, измерения интенсивности транспирации и непосредственные микроскопические наблюдения. Они подтвердили правильность теории сцепления. К началу 50-х годов удалось окончательно установить, что вода перемещается по неживым элементам ксилемы в направлении снижения гидростатического градиента, обусловленного транспирацией. Водные нити в сосудах находятся в состоянии натяжения, которое поддерживается силами сцепления между молекулами воды и силами прилипания воды к клеточным оболочкам.
Использование красящих веществ, радиоактивных изотопов и других индикаторов позволило определить скорость передвижения воды в растении. Для этого немецкий исследователь Б. Губер (1932) разработал специальный термоэлектрический метод, ранее использованный Г. Райном (1928) для определения скорости кровообращения. Согласно данным Губера, наименьшей скоростью отличаются хвойные, наибольшей - тропические лианы, травы и деревья. Измерения скорости тока в различных частях растений показали, что она меняется в зависимости от их обеспеченности водой, интенсивности транспирации и структурных особенностей растения.
Наибольшее количество работ по вопросам водного обмена относится к проблеме транспирации - процессу испарения воды растением. Процесс этот оказался очень интенсивным. По подсчетам Г. Шредера (1919), из всей солнечной энергии, затрачиваемой на процессы испарения на нашей планете, 2,5 - 5,0% падает на долю растений.
При изучении зависимости транспирации от внешних и внутренних факторов большинство исследователей концентрировало свое внимание на первых при различных экологических условиях (болото, поле, засушливые районы, освещенные и затененные участки тропического пояса и т. д.).
В 1900 г. английские ученые Г. Т. Броун и Ф. Эскомб, исследовав диффузию газов через перфорированные поверхности и через устьица, установили, что скорость диффузии зависит не от общей площади отверстий устьиц, а от их диаметра. Важная роль движения замыкающих клеток устьиц в регулировании транспирации подтверждена опытами. Однако до сих пор не раскрыты все закономерности, определяющие реакцию устьичных движений на транспирацию, зависящую от комплекса внутренних факторов.
Первые исследования в данном направлении провел Р. К. Найт (1916 - 1922). Изучая интенсивность транспирации в связи с функцией устьиц, он определил ее зависимость от содержания воды в клетках мезофилла и прежде всего от осмотических свойств последних. Исследования 30 - 40-х годов расширили проблему и позволили установить, что из числа внутренних факторов наибольшее влияние на транспирацию оказывают строение клеточных оболочек мезофилла, сопротивление движению воды, осмотические свойства клеток, величина внутренней испаряющей поверхности, число и размещение устьиц. В 30-е годы много внимания стали уделять также изучению действия на транспирацию болезней растений и различных химических средств борьбы с ними.
Особенно широко развернулось изучение водного режима растений с 40-х годов благодаря созданию новых методов исследования (изотопных индикаторов, радиоспектроскопии и др.). Они позволили развить ряд новых представлений о его регулировании как в самом растении на субклеточном уровне, так и в системе почва - растение - атмосфера. Развитию исследований по этому вопросу способствовали работы П. Д. Крамера (1949) и А. Крафтса с сотрудниками (1949). В нашей стране они велись под руководством Н. А. Максимова (1952). Существенно расширилось изучение взаимосвязи водного режима растений с фотосинтезом, дыханием, с аномальными отклонениями в обмене веществ, ведущими к изменению количества и состава содержащихся в растении соединений, с прорастанием семян, ростом растений и другими процессами. При помощи изотопной воды Г. Хюбнер (1960), А. Л. Курсанов и Б. Б. Вартапетян (1961) показали, что вся находящаяся в растении вода способна одинаково легко и быстро обмениваться с водой окружающей среды.
Новые методы позволили глубже изучить внутриклеточный водообмен. Ряд исследователей 30 - 40-х годов отстаивали мнение об "активной секреции" воды протоплазмой в вакуоль. А. Крафтс с сотрудниками (1951) объясняли этот процесс электроосмосом, возникающим в результате разности потенциалов по обе стороны протоплазматической мембраны, а А. М. Алексеев (1950) - тенденцией вещества к рассеиваемости. Позднее Алексеев (1960) развил представление о передвижении вод через мезоплазму как активированную диффузию, при которой молекулы воды должны преодолевать потенциальные энергетические барьеры. Работы 50 - 60-х годов преимущественно советских исследователей (К. С. Тринчер, 1966; А. М. Алексеев, 1969; и др.) показали, что водный режим растений должен изучаться не только на основе законов осмоса, но и с точки зрения коллоидной химии и биохимии.
Новые данные по этому вопросу заставили критически пересмотреть и представления о связывании внутриклеточной воды. Было установлено, что водный режим растений связан с коллоидальной структурой протоплазмы и ее высокомолекулярных компонентов, с количеством, строением и состоянием белковых веществ, с фосфорным и белковым обменами, обусловливающими энергетический уровень клетки и ее компонентов. Внесена ясность в вопрос о формах воды, среди которых различают "свободную", "осмотическисвязанную" и "коллоидносвязанную" (Н. А. Гусев, 1966).
Классификацию почвенной влаги на основании ее подвижности дали С. И. Долгов (1948) и А. А. Роде (1952). Особое внимание они уделили изучению доступных растению форм воды, значению подвижности почвенной воды и роста корней для водного снабжения растений. Все вопросы имели большое практическое значение, способствуя решению такой важной сельскохозяйственной проблемы, как рациональное использование осадков и орошения.
Физиология субклеточных структур
На X Международном ботаническом конгрессе в Эдинбурге (1964) особое внимание привлекла работа двух не существовавших на предыдущих конгрессах секций - физиологии растительной клетки и ее тонкой структуры и функции. Необходимость создания этих секций была вызвана значительными успехами и перспективностью исследований на клеточном и субклеточном уровнях, позволяющими проникать в тончайшие механизмы фотосинтеза, дыхания, поглощения веществ и других процессов. Эти успехи стали возможны благодаря существенному прогрессу методов исследования клетки и ее физиологии (см. главы. 10 и 11). Следует особо отметить приемы К. Мюлеталера и его сотрудников (1964), которые во избежание артефактных изменений тонкой структуры клеток заменили химическую фиксацию материала его быстрым замораживанием. Этот прием позволил обнаружить ряд деталей в структуре цитоплазматической мембраны, ядерной оболочки, квантосомы в ламмелах хлоропластов, оксиомы в митохондриях и другие органеллы, ускользавшие от наблюдения при обычных методах фиксации.
Получение чистых фракций неповрежденных клеточных органелл позволило уже в начале 40-х годов приступить к более детальному изучению не только структуры, но и функции растительных митохондрий. Согласно многолетним исследованиям американского биохимика Д. Э. Грина (1963), именно с этими клеточными органеллами связаны три важнейших процесса: окисление в цикле трикарбоновых кислот, а вне его - аминокислот, жирных кислот и других соединений; перенос электронов и оксислительное фосфорилирование (см. главу 6). Сравнение митохондрий растительных и животных клеток и характера протекающих в них процессов обнаружило почти полную аналогию их функций.
В 50-х годах в протоплазме растительных клеток были обнаружены рибосомы - рибонуклеопротеидные частицы, впервые наиболее подробно описанные П. Тсо с сотрудниками (1956 - 1958). Тогда же было установлено, что с деятельностью этих частиц как в животной, так и в растительной клетке связан синтез белка.
В настоящее время ведутся интенсивные исследования механизмов на отдельных этапах белкового синтеза и участия рибосом в этих процессах (см. главу 23).
Важные данные получены в 60-х годах о структуре и функции хлоропластов. Электронно-микроскопические наблюдения и исследования биохимического состава и ферментативной активности этих органелл позволили установить наличие у них ламеллярной (пластинчатой) структуры. Ламеллы состоят из тонких чередующихся друг с другом слоев белка и липоидов. Через определенные промежутки несколько ламелл соединяются между собой, образуя в местах соединения уплотненные плоские участки - граны. Каждый слой ламелл в свою очередь состоит из плотно прилегающих частиц, внешне напоминающих зернистую поверхность. Впервые эти частицы были обнаружены на электронных микрофотографиях Э. Штейнмана (1955). М. Кальвин (1962) назвал их квантосомами.
В хлоропластах уже к началу 50-х годов были найдены почти все ферменты, участвующие в метаболизме растений, что позволило Н. М. Сисакяну (1954) характеризовать эти пластиды как "депо энзимов". Наибольшее внимание было уделено изучению пиридиннуклеотидредуктазы, трансгидрогеназы и ферментам фосфорного обмена - АТФазы, фосфатаз и др.
Важнейшим событием было обнаружение в конце 50-х - начале 60-х годов в хлоропластах рибосом, что окончательно утвердило мнение о возможности синтеза белка в хлоропластах (П. Тсо, 1958; В. Литлтон, 1962; Г. Брауерман, 1963; и др.).
До начала 40-х годов считалось общепризнанным, что процессы дыхания, фотосинтеза и ряд других биохимических реакций неразрывно связаны с растительной клеткой как целостной структурной единицей. Опыты Р. Хилла (1939), а затем А. Е. Бойченко (1949), Д. И. Арнона (1954, 1962) и других опровергли это мнение и в отношении хлоропластов. Исследования фрагментов этих структур привели в выводу, что даже их мельчайшие частицы содержат все фотосинтетические пигменты в таком же соотношении, как и неповрежденные клетки; передача энергии в таких фрагментах идет так же, как и в целых клетках, хотя с меньшей интенсивностью.
Новые данные получены также о структуре и функции ядра, ядерных образований и оболочки. Американские биохимики А. Е. Мирский (1963) и В. Г. Олфри (1963) обнаружили существование в изолированных ядрах процессов фосфорилирования и синтеза АТФ, доказав тем самым, что митохондрии не являются единственными структурами, в которых возможны эти процессы. В 1965 г. Дж. Боннеру удалось выделить из клеток ростков гороха хромосомы и показать, что в присутствии рибосом, аминокислот и рибозодифосфатов они и в изолированном состоянии могут синтезировать белки типа глобулинов, свойственные только этой стадии развития гороха. Электронно-микроскопические исследования позволили установить, что ядерная оболочка состоит из двух слоев - внутреннего, гомогенного, и наружного, пронизанного порами. Последние обеспечивают возможность беспрепятственного обмена веществ между ядром и цитоплазмой (Г. Г. Галлан, 1952).
В изучении тончайших структур и функций клеточных органелл заметную роль начинают играть работы, в которых структура и функция рассматриваются в развитии. Современные цитологические исследования ведут к расшифровке физиологических процессов, протекающих во вновь открываемых субклеточных элементах. Именно эта область исследований служит наиболее ярким примером того, как все более стираются границы между биохимией, цитологией и физиологией растений и как постепенно обрисовываются контуры синтеза этих наук на более высоком уровне.
Рост и ростовые процессы
В XX в. исследования роста и ростовых процессов приобретали все более ясно выраженную биохимическую направленность: прорастающие семена изучались с точки зрения активности различных ферментов, особенно дыхательных; ростовые процессы клетки рассматривались в тесной связи с метаболизмом не только самой клетки, но и всего растения в целом и т. д.
Большая часть исследований относилась к изучению различных проявлений ростовых движений - тропизмов (фото-, гео-, гидро-, травма-, и гаптотропизмов) и настий. Кроме этих типов движений В. Н. Половцева (1909) обнаружила существование у растений аэротропизма - движения побегов и корней в направлении поступления кислорода, а Ф. М. Породко (1915) - хемотропизма - изменения направления роста при одностороннем действии химических раздражителей. Особое внимание привлекло изучение причин этих явлений.
Для объяснения геотропизма Г. Габерландт (1900 - 1908) и Б. Немец (1901) развили так называемую статолитную теорию, согласно которой у высших растений имеются особые органы восприятия силы тяжести, подобные отолитам животных. Аналогичных представлений Габерландт (1905) придерживался и в отношении фототропизма. Недостаточное знание биохимии ростовых процессов породило ряд механистических толкований роста, окончательный удар которым был нанесен утверждением гормональной теории тропизма, направившей изучение вопроса в новое русло - глубокого физиологического анализа этих явлений.
Предположения о существовании особых ростовых веществ в растении, высказывавшиеся в прошлом столетии, в начале XX в. получили экспериментальное подтверждение. Датский исследователь П. Бойсен-Йенсен (1910 - 1911) доказал наличие в растении гормона роста. Опыты А. Пааля (1914, 1918), показавшие, что передача раздражения не связана с электрическими явлениями, а также П. Штарка (1921, 1922), обратившего внимание на необходимость химического исследования этого явления, не оставляли сомнений, что явление тропизмов обусловлено действием какого-то вещества. К началу XX в. относятся первые исследования Н. Г. Холодным геотропизма (1906) и хемотропизма (1908) корней, на основе которых он развил ионную теорию геотропизма. В 1926 г. Холодный разработал стройное представление о механизме геотропизма и фототропизма. Одновременно и независимо от него Ф. В. Вент (1926, 1928) выдвинул теорию, согласно которой причиной фототропической реакции растений является отклонение тока гормона роста к затененной стороне органа.
Дальнейшему совершенствованию теории Холодного - Вента способствовали выделение и химическое изучение ростовых веществ. Вплотную к получению ростового гормона подошли Г. Е. Дольк (1929) и Ф. А. Вент (1934). Им удалось добиться диффузии этого вещества из растительного сока в агар-агар и предпринять первые попытки его качественного и количественного изучения. Исследованиями физиологов заинтересовались химики. Немецкому биохимику Ф. Кёглю с сотрудниками (1931 - 1935) удалось выделить фитогормон в химически чистом виде как из растений, так и из животных, в организм которых они попадают с растительной пищей. Кёгль называл эти вещества ауксинами. В 1934 г. Ф. Кёгль, А. Гааген-Смит и X. Эркслебен сообщили, что получили из различных плесневых грибов, мочи животных, а позднее и из высших растений еще одно ростовое вещество - [J-индолилуксусную кислоту, физиологическая активность которой почти вдвое ниже активности ауксинов. Они назвали его гетероауксином. Холодный (1931 - 1937) и Вент (1932) первыми стали исследовать механизм перемещения ауксинов в растении.
С изучением ауксинов, стимулирующих рост клетки в фазе растяжения, было тесно связано исследование дополнительных факторов роста - стимуляторов деления клетки. В 1901 г. Е. Вильде открыл в солоде вещество, ускоряющее размножение дрожжевых клеток, и назвал его биосом. Химическая природа биоса была выяснена лишь три десятилетия спустя. Ф. Кёгль (1932 - 1936) и В. В. Вильямс (1933 - 1935) установили, что биос представляет собой комплекс нескольких соединений, являющихся компонентами витаминов группы В. Было обнаружено, что вещества этого комплекса синтезируются в листьях, а оттуда поступают в различные ткани растений, в частности в точки роста.
В 1908 г. А. Н. Набоких высказал предположение о выделении "возбудителей" роста пораненными тканями растений, а в 1913 г. Г. Габерландт открыл некрогормоны (раневые гормоны) - вещества, образующиеся при распаде поврежденных тканей и способные вызывать деление покоящихся клеток высших растений в месте поранения. В 1926 г. японский биохимик Е. Куросава экстрагировал из почвенного гриба гибберелла, паразитирующего на рисе, сок, содержащий вещество высокой физиологической активности, которое назвали гиббереллином. Благодаря работам японских, американских и английских исследователей в 40 - 50-х годах удалось обнаружить гиббереллины также в клетках высших растений, выделить их и изучить стимулирующее действие на рост, накопление сухой массы и ускорение развития многих растений. В настоящее время известно более 30 различных веществ такого типа, относящихся к алициклическим карбоновым кислотам. К. О. Миллер (1955) извлек из спермы сельди кинин - вещество, сильно ускоряющее деление клеток табака и разрастание опухолей. Выяснив, что кинин является производным аденина (постоянной составной части нуклеиновых кислот), он установил связь между делением клеток и их нуклеиновым обменом.
Открытие ауксинов и других стимуляторов роста дало возможность управлять этими процессами. Если прежде выведение почек растений из состояния глубокого покоя производилось охлаждением, действием паров эфира, теплыми ваннами, то начиная с 30-х годов для ускорения прорастания спящих почек, глазков у картофеля, стимуляции укоренения черенков стали использовать природные и синтетические ауксины. Опрыскивая водным раствором ауксина неопыленные завязи томатов, винограда и инжира, Ф. Густафсон (1936) и другие получили из них бессемянные (партенокарпические), но совершенно зрелые плоды.
Б. П. Токин в 1928 г. в докладе, а в 1930 г.- в печати сообщил об открытии им у высших растений защитных веществ, способных подавлять рост бактерий, грибов и простейших. Эти вещества были названы фитонцидами. В книге "Бактерициды растительного происхождения (фитонциды)" (1942 и в последующих изданиях) Токин изложил результаты изучения их свойств, химической природы, мощности и диапазона действия, влияния на состав биоценозов, а также указал на перспективы их использования в медицине, сельском хозяйстве и пищевой промышленности. Открытие фитонцидов положило начало широким поискам антибиотиков, выделяемых высшими растениями.
В течение последних десятилетий изучение ростовых процессов шло по пути раскрытия внутренних условий роста и установления связи ростовых процессов с физиологически активными веществами - гормонами роста, особенно гетероауксином и кининами. Было также установлено, что некоторые вещества, не встречающиеся в растениях, могут оказывать сильное влияние на ростовые и формообразовательные процессы. Такие вещества получили название стимуляторов роста. К ним относятся 2,4-дихлорфеноксиуксусная, а-нафтилмасляная и другие кислоты. Взятые в небольших концентрациях, они так же, как естественный гормон роста гетероауксин, способствуют корнеобразованию, почему получили широкое применение в практике размножения растений черенками.
В 50 - 60-е годы предметом интенсивного исследования стали природные (абсцизовая кислота, кумарин, флоридзин) и синтетические (гидразид малеиновой кислоты, трийодбензойная кислота, хлорхолинхлорид, морфактинхлорфлуоренол) ингибиторы роста растений. Природные ингибиторы роста играют существенную роль в состоянии покоя, а также при воздействии неблагоприятных факторов среды, например засухи. Ингибиторы находят применение в сельскохозяйственной практике. Их используют, например, для задержки прорастания клубней картофеля.
Было установлено, что физиологически активные вещества оказывают прямой положительный эффект только в малых дозах. Взятые в более высоких концентрациях, они вызывают отравление растений и применяются в качестве гербицидов для уничтожения сорняков в посевах. Таковы гербициды избирательного действия, например 2,4-дихлорфеноксиуксусная кислота. Как показали работы И. И. Гунара (1952), гербициды приводят к нарушению азотного и углеводного обмена веществ, а также фотосинтеза, дыхания и других процессов. Впрочем, и вредное действие веществ может иногда оказаться хозяйственно полезным. Так, для механизированной уборки хлопчатника большое значение имеют и дефолиация (удаление листьев), я десикация (высыхание листьев).
В 60-х годах П. А. Генкель показал, что интенсивный рост стебля и корня связан с усиленным обменом веществ и ростом у них той стороны, где скапливается ауксин. По данным Л. Браунера (1959), под действием земного тяготения возникает "геоэлектрический эффект", т. е. изменения в биоэлектрических явлениях. Аналогичная поляризация органа, подвергавшегося раздражению, наблюдалась Браунером также и при одностороннем освещении, причем возбужденная сторона становится электроотрицательной.
Помимо ростовых движений у растений стали различать движения сократительные, например складывание листочков у стыдливой мимозы (Mimosa pudica) и у кислицы (Oxalis acetosella). В. Н. Жолкевич и А. Г. Четвериков (1967) показали, что эти движения связаны с изменениями свободных радикалов, т. е. с превращениями энергии в растении. Б. Ф. Поглазов (1965) обнаружил, что у тропического растения Desmodium gyrans наблюдается высокая АТФазная активность, тогда как у растений, не способных к движению, она незначительна. Работы Поглазова сделали весьма вероятным предположение, что многие движения "сна" являются не ростовыми, а сократительными.
Давно отмечено, что ростовые процессы и движения растений совершаются ритмически, следуя обычным периодическим явлениям - смене дня и ночи и времен года ("физиологические часы"). Однако в ряде случаев связи между ритмом того или иного физиологического процесса и окружающими условиями обнаружить не удается. Тогда говорят об эндогенных ритмах растения.
Устойчивость растений к неблагоприятным условиям
В 20 - 30-х годах XX в. преимущественно благодаря работам советских физиологов возникло новое направление исследований - устойчивости растений к неблагоприятным условиям.
Наиболее интенсивно изучалась засухоустойчивость растений, точнее те свойства, которые позволяют им переносить обезвоживание и перегрев. Для правильного понимания природы этого явления немаловажное значение имели исследования В. Р. Заленского (1904), показавшего, что листья верхних ярусов отличаются более ксероморфным строением, так как развиваются в условиях большей физиологической сухости. Значительной популярностью до 1916 г. пользовалась точка зрения А. Ф. Шимпера, который объяснял засухоустойчивость растений способностью экономно расходовать воду. Несостоятельность такого мнения впервые показали американские исследователи Л. Бригтс и Г. Шанц (1911 - 1913), проводившие свои опыты в засушливой зоне штата Колорадо (США), а затем Н. А. Максимов (1914 - 1916), создавший новую теорию засухоустойчивости. Исследования Максимова положили начало пониманию засухоустойчивости как приспособления, позволяющего растению переносить сильное завядание с наименьшим ущербом как для отдельного индивидуума, так и для вида в целом. Вслед за И. И. Тумановым (1926, 1930), обнаружившим способность растений к закаливанию, П. А. Генкель (1946) разработал метод предпосевного закаливания семян, существенно повышающий устойчивость растений к засухе без снижения урожайности.
Многие советские фитофизиологи показали, что физиологическая основа засухоустойчивости у разных групп растений различна и что она зависит от условий, в которых шло их формирование. Она характеризуется теми изменениями в обмене веществ, которые возникают при обезвоживании. Н. М. Сисакян (1940) установил, что засухоустойчивость обусловлена высокой синтетической способностью растения и что его повреждение и гибель происходят в результате падения содержания белковых веществ в протопласте ниже определенной нормы.
В 60-е годы исследователи обратили особое внимание на изучение зависимости устойчивости растений от генетических факторов. Н. А. Сатарова и Е. К. Творус (1965) установили, что у наиболее устойчивых растений и растений, семена которых прошли предпосевную закалку, распада белков с образованием аммиака не происходит, а их ферментативные системы в условиях засухи оказываются более стабильными. При этом в них происходит обновление белка, а после засухи - новообразование белковых веществ и репарация. Б. Кесслер (1961) выяснил значение в выработке засухоустойчивости нуклеиновых кислот, в частности, РНК. Благодаря электронной микроскопии стало возможным изучение изменений, происходящих в клеточных органеллах - рибосомах, полирибосомах и митохондриях - под влиянием засухи (Н. А. Сатарова, Е. К. Творус, 1965; и др.). В. Н. Жолкевич (1957, 1958) и Т. Ф. Корецкая (1959) показали, что под влиянием засухи происходит разобщение окисления и фосфорилирования в процессе дыхания и последнее становится энергетически неполноценным.
Как показали А. М. Алексеев и Н. А. Гусев (1957), под влиянием засухи в растении происходит изменение структурного состояния воды, что влечет за собой глубокие изменения в макромолекулах белков и нарушения течения биохимических процессов.
Было выяснено, что по мере развития растения его засухоустойчивость обычно повышается, но этот процесс идет лишь до момента образования генеративных органов. Особенно чувствительной к засухе оказалась пыльца. Исследователи пришли к выводу, что в условиях засухи важным фактором нормального образования генеративных элементов является предпосевное закаливание.
В настоящее время во многом выяснена природа ряда ксерофитов и пойкилоксерофитов. Так, например, установлено, что жароустойчивость таких суккулентов, как кактусы, агавы, алоэ, обусловлена большой вязкостью протоплазмы и высоким содержанием связанной воды. Зато они не способны выносить обезвоживание, поскольку обладают малой эластичностью протоплазмы. Кроме того, для обводнения биополимеров кутикулы эти растения используют метаболическую воду дыхания.
Принято считать, то пойкилоксерофиты впадают в настоящий анабиоз и могут безгранично долго сохранять жизнеспособность. Так, Р. Е. Камерон (1962) обнаружил, что синезеленая водоросль Stratonostoc commune в гербарии сохранила свою жизнеспособность в течение 107 лет, а семена лотоса, по данным Ога [цит. по Г. Молишу (1933) ], не утратили ее, пролежав в торфяном болоте свыше 1000 лет. Одни исследователи считают, что во время анабиоза жизнь прерывается (П. Ю. Шмидт, 195,5), другие рассматривают его как период скрытой жизни (А. И. Опарин, 1960).
К числу неблагоприятных факторов, связанных с недостаточной увлажненностью, относится повышенная соленость почв. Уже в начале нашего века было выяснено, что засоленные почвы образуются в засушливых районах, где редко выпадающие осадки не в состоянии вымыть из почвы накапливающиеся при выветривании минеральные соли. Изучение произрастающих на таких почвах солеустойчивых растений (галофитов) провел советский ботаник Б. А. Келлер (1923, 1927). Он обнаружил, что для солянок, растущих на мокрых солончаках, хлористый натрий не является физиологически необходимым. О. Штокер (1928) показал, что суккулентность галофитов иная, чем у ксерофитных суккулентов, и она не связана с замедленной отдачей воды. Позднее П. А. Генкель и А. А. Шахов (1945) и А. А. Шахов (1956) доказали, что для галофитов солончаки не являются физиологически сухими в связи с близостью залегания грунтовых вод, а также благодаря высокому осмотическому давлению, позволяющему им развивать большую сосущую силу и беспрепятственно расти на высоком фоне засоления. Исследователи установили также происхождение галофитов от гликофитов. Теперь окончательно доказано, что при высокой засоленности почв на растение оказывают вредное действие прежде всего соли, а уже потом сказывается отрицательное значение осмотического давления почвенного раствора.
М. Д. Кушниренко (1950) показала, что одни и те же галофиты неодинаково адаптируются на различных солончаках: на сульфатных их интенсивность обмена повышается, а на хлоридных, наоборот, понижается. Б. П. Строгонов (1962) установил, что в основе этого явления лежат разнокачественные реакции на почвы с хлоридным, сульфатным и карбонатным засолением. Выяснение разнокачественности засоления почв поставило вопрос о необходимости соответственно дифференцированной агротехники и селекции. Были изучены и некоторые общие реакции растений на засоление.
Предметом исследований явился также вопрос о характере токсического действия солей на растения. Было выявлено различное действие высоких концентраций ряда солей на субмикроскопическую структуру хлоропластов кукурузы. Косвенное их влияние проявлялось в образовании ядовитых соединений и отравлении растений. В. А. Соловьев (1960) пришел к выводу, что засоление нарушает работу трансаминаз и процесс переаминирования, в результате чего накапливается ряд ядовитых для растений промежуточных продуктов обмена. Согласно Б. Кесслеру (1964), засоление затрагивает и другие системы, в частности, нуклеиновый обмен. В условиях засоления снижается отношение ДНК/РНК и сильно угнетается образование РНК, а при адаптации к засолению возрастает устойчивость рибосом, связанных с синтезом белка. Исследовалось также влияние засоления на углеводный обмен, дыхание и фотосинтез.
Одновременно с изучением действия солей на растение шла разработка методов повышения солеустойчивости. Одним из них явился метод предпосевного солевого закаливания, предложенный П. А. Генкелем (1940). В 50 - 60-х годах Генкель с сотрудниками разработали методы солевого закаливания применительно к хлоридному, сульфатному и карбонатному засолению почвы. Метод повышения солеустойчивости в ряде вариантов был испытан в работах большого числа авторов применительно к хлопчатнику, сахарной свекле, к чубукам винограда, сое и чумизе, просу и пшенице.
Способность растения, именуемая зимостойкостью, включает в себя такие явления, как морозостойкость, холодостойкость, а также устойчивость к зимней засухе, выпреванию, вымоканию и т. п. Определяющим моментом зимостойкости является морозоустойчивость, т. е. способность растений противостоять температурам ниже 0°. Согласно первой теории морозостойкости (Г. Мюллер-Тургау, 1880, 1886; Г. Молиш, 1897), гибель растений при низких температурах происходит в результате обезвоживания протоплазмы, как следствие образования в межклетниках кристаллов льда. Против этой теории выступил К. Мец (1905), который не только отрицал вредное действие льдообразования, но, наоборот, считал этот процесс полезным.
Необоснованность представлений Меца была доказана в результате исследований Н. А. Максимова (1913 - 1917). Обратив особое внимание на биохимические изменения, происходящие в растении осенью и зимой, Максимов показал, что их вымерзание - результат полного нарушения структуры протоплазмы вследствие обезвоживания и механического давления льда. Растения гибнут не от низкой температуры, как таковой, а от кристаллизации воды. Кристаллы льда подсушивают протоплазму, вызывая коагуляцию ее коллоидов, что ведет к гибели клетки. Если введением концентрированных растворов Сахаров уменьшить или совсем предотвратить образование кристаллов льда, то ткани растений могут оставаться длительное время жизнеспособными и в условиях низких температур. П. Л. Богданов (1931) и Б. С. Мошков (1935) установили большое значение осеннего укорочения дня для перезимовки растений. Наибольших успехов в изучении приспособления (закаливания) растений к низким температурам достиг И. И. Туманов (1951). Согласно Туманову, морозостойкость растений вырабатывается в ходе двух стадий закаливания - осенью и в начале зимы. На обеих стадиях происходят определенные изменения в структуре и составе протоплазмы клеток, повышающие их зимостойкость. М. Ф. Бугаевский (1940) изучил последовательность льдообразования при замерзании молодых тканей пшеницы, а Т. С. Сулакадзе (1945) показала, что клетки закаленных растений более устойчивы к действию кристаллов льда, чем незакаленных. Многочисленными исследованиями было установлено, что одним из важнейших моментов закаливания растения является прекращение его роста и переход в состояние покоя.
Дальнейшее изучение процесса закаливания растений показало, что во время морозов растения повреждаются и гибнут из-за накопления льда не в межклетниках, а в самой протоплазме, где он образуется при гораздо более низких температурах. Г. А. Самыгин (1960) выявил разные типы льдообразования в зависимости от скорости охлаждения. Повышению устойчивости растений к морозу способствует более короткий день, когда количество гормонов роста в растении резко снижается, а содержание ингибиторов роста, наоборот, возрастает, когда падает интенсивность обмена веществ и изменяется белковый состав. М. М. Окунцов и О. Ф. Аксенова (1960) обнаружили у озимой пшеницы глубокую перестройку дыхательных систем.
Новое объяснение природы морозоустойчивости растений дал Дж. Левитт (1962). По его мнению, определяющую роль в выработке устойчивости к низким температурам играют сульфгидрильные группы белков протоплазмы, тогда как дисульфидные группы тормозят этот процесс. При быстром оттаивании льда происходит заполнение водой межмицеллярного пространства и ее гидростатическое давление вызывает разрыв образовавшейся между молекулами белка водородной связи, обладающей меньшей энергией по сравнению с дисульфидной. Концепция Левитта интересна как первая попытка объединить теорию механического повреждения кристаллами льда с биохимическим направлением в исследовании морозоустойчивости.
П. А. Генкель и Е. 3. Окнина (1948, 1964 и позднее) развили представление об "обособлении" протоплазмы, согласно которому растение, готовящееся к перезимовке, впадает в состояние глубокого покоя; при этом плазмодесмы разобщаются и образующийся в межклетниках лед не может повредить протоплазму. Явление обособления протоплазмы было подтверждено многими исследователями как в нашей стране, так и за рубежом. На основании концепции обособления были предложены методы диагностики состояния покоя и морозоустойчивости плодовых растений, которые используются многими плодоводами-оригинаторами при выведении новых сортов.
Исследованиями П. А. Генкеля и Е. 3. Окниной (1964), И. И. Туманова (1967) и других были выявлены три фазы покоя - органический, глубокий и вынужденный. Согласно взглядам Л. И. Сергеева (1960 и позднее), важной биологической характеристикой древесных пород является их годичный морфофизиологический ритм. В периоды наиболее интенсивного метаболизма устойчивость структуры их протоплазмы понижается, а при падении метаболизма, наоборот, повышается. При глубоком (органическом) покое происходит отток ряда соединений из листьев в зимующие органы, где идет синтез РНК, белков, липоидов и олигосахаридов. Одной из причин глубокого покоя является низкий уровень РНК и особенно ДНК на эмбриональной стадии почек.
Большой интерес представляют опыты японского ученого А. Сакаи (1956 - 1961), а также И. И. Туманова и О. А. Красавцева (1962), изучавших возможности достижения безграничной морозоустойчивости у древесных пород. Еще в 1938 г. американский исследователь Б. И. Лейет открыл явление витрификации протоплазмы, заключающееся в быстром замерзании содержащейся в ней воды (при температуре ниже - 150°) без образования кристаллов льда (или очень мелких кристаллов). Возникающий в этом случае аморфный лед не повреждает протоплазму, и после ее быстрого оттаивания (девитрификации) растение возвращается к жизни. В опытах Туманова и Красавцева (1959) закаленные ветки смородины, сосны и березы при ступенчатом охлаждении оставались живыми при температуре до - 195°. Большой вклад в изучение морозоустойчивости плодовых культур и ее диагностику внесли работы М. А. Соловьевой (1967).
Среди методов диагностики наиболее надежными считаются многочисленные способы промораживания в холодильных камерах и фитотронах, а также в полевых холодильных установках.
В последние два десятилетия большое внимание стали уделять также проблеме холодостойкости - способности растений переносить без повреждений низкие положительные температуры, близкие к 0°. Еще Г. Молиш (1897) высказывал правильное мнение, что в условиях пониженных температур у растений тропического и субтропического происхождения нарушается обмен веществ. В 50 - 60-х годах оно получило развитие в работах П. А. Генкеля, К. П. Марголиной и В. Н. Жолкевича. Л. А. Незговоров и А. К. Соловьев (1958) показали зависимость холодостойкости растений от патогенной микрофлоры почвы.
В направлении повышения холодостойкости теплолюбивых растений методом закаливания семян работали В. И. Эдельштейн (1944, 1962), А. Е. Воронова (1950, 1953, 19,59) и др. Изучение происходящих при этом физиологических изменений шло по двум путям: с одной стороны, исследовали стимулирующее действие переменных температур и их влияние на рост и развитие растений, с другой - основное внимание концентрировали на холодостойкости растений. В результате было выявлено значение пониженной вязкости протоплазмы и усиления обмена веществ для повышения холодостойкости теплолюбивых растений. Закаленные переменными температурами растения сохраняли энергетически более полноценное дыхание, более стабильные фотосинтез и активность ряда ферментов и другие физиологические процессы и при воздействии низких положительных температур.
Прямое отношение к проблеме холодостойкости имело открытие в 40-х годах А. В. Благовещенским и В. П. Филатовым так называемых биогенных стимуляторов - веществ, в норме не свойственных живым тканям растений или животных и возникающих в них при низкой положительной температуре (около +1°) в качестве защитного средства. Благовещенский (1949 - 1950) показал, что в малых количествах эти чуждые организму вещества оказывают стимулирующее, а в больших - отравляющее действие на плазму.
В последнее время возрос интерес к влиянию на растения заморозков, вызывающих образование кристаллов льда в межклетниках. Работы С. Н. Дроздова (1968) показали, что под действием заморозков в клетках происходят структурные изменения белков и нарушается нормальный синтез АТФ.
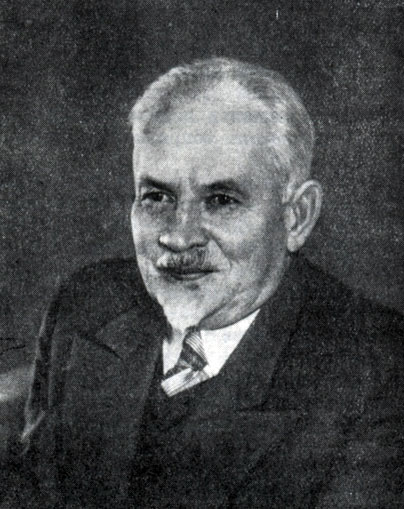
Николай Александрович Максимов (1880 - 1952)
По данным А. И. Коровина и его сотрудников (1968), после заморозка содержание органических форм фосфора и азота, в частности макроэргического фосфора и РНК, сильно снижается, а неорганических, наоборот, резко возрастает (накапливаются амиды и аммиак). Изменяя уровень минерального питания путем повышения концентрации различных элементов, можно влиять на морозоустойчивость.
Экологическая физиология растений
В 50-х годах в СССР складывается новый - физиологический - подход к изучению устойчивости растений и возникает новое направление исследований - экологическая физиология растений. Основополагающее значение для ее создания имели труды Н. А. Максимова* (1913, 1926, 1952; Ленинская премия, 1930), подходившего к изучению явлений устойчивости растений с физиологических позиций. Такой подход позволил исследовать устойчивость не только на организменном и микроскопическом, но в последнее время также и на субмикроскопическом и даже молекулярном уровнях. Он дал, кроме того, возможность установить связь устойчивости с такими общебиологическими явлениями, как анабиоз, передача энергии при засухе, адаптационный процесс, идущий под влиянием окружающих условий, и т. д.
* (Подробнее см.: П. А. Генкель. Научная деятельность Николая Александровича Максимова и его роль в создании экологической физиологии растений.- В сб. "Проблемы физиологии растений (исторические очерки)". "Наука", 1969, стр. 306 - 331.)
Одни исследователи (Д. Н. Насонов, В. Я. Александров, 1940) полагают, что адаптационный процесс имеет неспецифический характер, другие (П. А. Генкель, 1960, 1967) стоят на точке зрения его специфичности. Согласно последней точке зрения, влияние неблагоприятных условий (холода, мороза, засухи, засоления) вызывает у растений комплекс ответных защитноприспособительных реакций, одна часть которых носит общий неспецифический характер, а другая вполне специфична. В то же время, сравнение различных видов устойчивости позволило в некоторых случаях установить наличие между ними определенной связи, которая получила название сопряженной устойчивости (П. А. Генкель, С. В. Кушниренко, 1966).
Что касается вопроса об адаптации к отдельным неблагоприятным факторам, то наиболее обоснованными можно считать представления О. Штокера и X. Росса (1956, 1967) о фазах реакции и реституции (восстановления), наблюдающихся у растения в ответ на засуху. Различение этих фаз, в которых находится растение, по мнению Штокера, очень важно для оценки его состояния.
На современном этапе развития физиологии растений отчетливо проявились две основные тенденции, характерные для всей биологии последних трех десятилетий,- ведение исследований в направлениях редукционизма и интегратизма. При этом следует заметить, что первое из них - изучение процессов жизнедеятельности растений на молекулярном уровне - по объему работ несколько уступает второму направлению - интеграционным исследованиям взаимодействия процессов в растениях на разных уровнях познания (от молекулярного до организменного). Это объясняется спецификой фитофизиологии, в центре внимания которой стоит растительный организм как целое, в отличие от биохимии растений, где преобладают работы первого направления. Пока нет оснований предполагать, что существующее соотношение двух названных направлений в фитофизиологических исследованиях в дальнейшем изменится. Несомненно, однако, что со временем гораздо шире развернется изучение взаимосвязей между отдельными растениями, составляющими фитоценоз и, в частности, агроценоз как наиболее просто устроенную экосистему.
Фитофизиологи всегда помнили заветную мечту К. А. Тимирязева "подчинить окружающую природу разумной воле человека", являющуюся по существу целью всех биологических исследований. Теоретические работы по физиологии растений немало дали практике народного хозяйства и прежде всего сельскому хозяйству. В настоящее время физиологи и биохимики растений уделяют гораздо больше внимания разработке фундаментальных проблем науки. Это позволяет решать вопросы практики растениеводства на более высоком уровне.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'