4.8.3. Класс Oligochaeta (малощетинковые)
Дождевой червь (himbricus) - животное с длинным цилиндрическим телом, достигающим 12-18 см. Передний его конец конусообразный, а задний сплющен в дорсовентральном направлении. И хотя он обитает только на суше, до конца приспособиться к наземному образу жизни он не смог. Большую часть суток червь проводит под землей, зарывшись во влажную почву, спасаясь таким образом от высыхания. Он покидает свою нору лишь ночью, отправляясь на поиски пищи или полового партнера. Различия в строении тела нереиса и дождевого червя можно объяснить приспособлением последнего к наземному образу жизни.
Тело дождевого червя имеет обтекаемую форму и лишено каких-либо выростов, поскольку выросты могут препятствовать свободному продвижению червя в почве. Простомиум небольшой и округлый, чувствительных придатков нет. Все сегменты, за исключением первого и последнего несут по четыре пары щетинок: две брюшные и две вентро-латеральные. Щетинки выходят из расположенных в стенке тела щетинковых мешков. Специальные мышцы (ретракторы) могут втягивать щетинку внутрь или, напротив, выставлять ее наружу (протракторы) (рис. 4.29). Щетинки олигохет участвуют в локомоторной активности. Длинные щетинки, расположенные на 10-15, 26 и 32-37 сегментах, играют важную роль в процессе копуляции. Еще одна структура, используемая в процессе копуляции, - поясок, который находится на 32-37 сегментах (рис. 4.30). Эпителий пояска содержит множество железистых клеток, образуя на спинной и боковых поверхностях заметное утолщение, напоминающее седло. Поясок участвует в копуляции и формировании кокона.
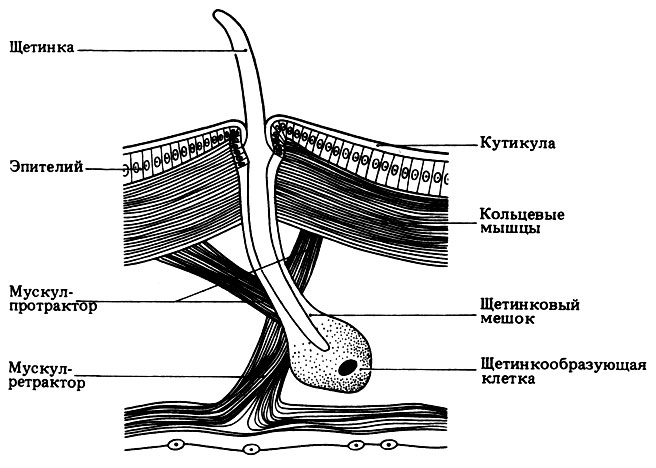
Рис. 4.29. Строение стенки тела дождевого червя (Lumbhcus terrestris). Поперечный разрез через область щетинки
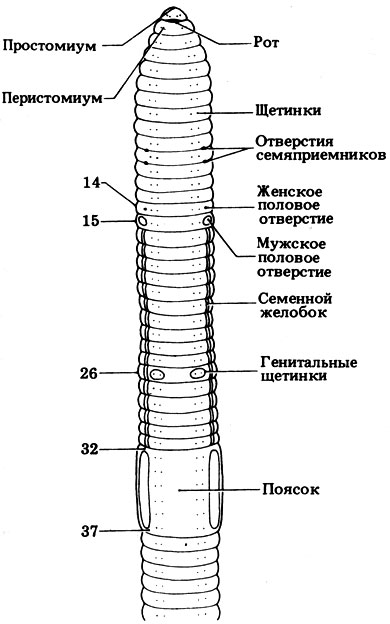
Рис. 4.30. Передний конец тела дождевого червя (Lumbricus terresths). Вид снизу
Строение стенки тела дождевого червя (рис. 4.29) сходно со строением стенки тела нереиса. Ротовое и анальное отверстия расположены на разных концах тела. Пища заглатывается в результате сокращения мышц неподвижной глотки. Кишечник прямой. Тифлозоль (продольная складка на спинной стороне кишки, вдающаяся в просвет кишечника) увеличивает площадь поверхности, участвующей в переваривании пищи и всасывании. Червь питается детритом; он заглатывает землю и переваривает содержащиеся в ней органические остатки. Питательные вещества, всасываясь, поступают в капилляры, окружающие стенку кишки. Основная часть почвы проходит через кишечник червя и выделяется в виде копролитов.
Тонкая кутикула олигохет увлажняется за счет постоянно выделяющейся через спинные поры целомической жидкости и слизи, секретируемой железистыми клетками эпителия. Именно через кутикулу происходит газообмен путем диффузии, и разветвленная сеть капилляров, расположенных в эпителии, обеспечивает этот процесс.
Все сегменты, за исключением трех первых и одного последнего, несут по паре нефридиев, выполняющих функции выделения и осморегуляции. Нефридии открываются на поверхности тела специальными отверстиями, расположенными несколько впереди от вентро-латеральных щетинок. Окружающие кишку хлорагогенные клетки также участвуют в процессах выделения.
Кровь от отдельных сегментов собирается в пульсирующий спинной сосуд. Пятью парами мускульных боковых "сердец", расположенных в 7-11 сегментах, она проталкивается в срединный брюшной сосуд. В "сердцах" и спинном сосуде имеются клапаны, препятствующие обратному току крови. От брюшного сосуда отходят боковые ветви, снабжающие кровью все сегменты тела. У дождевых червей не происходит заметной концентрации чувствительных структур на переднем конце тела. У них есть отдельные фоточувствительные клетки, а также клетки, реагирующие на химические вещества и свет; все эти клетки разбросаны в эпителии. Центральная нервная система дождевого червя сходна с нервной системой нереиса. В брюшной нервной цепочке у них имеются гигантские нервные волокна, которые в ответ на какое-либо сильное раздражение вызывают сокращение всей мускулатуры червя, хотя общая организация нервной системы может обеспечить координированную работу мускульных слоев, связанную с роющей или локомоторной активностью животного.
У дождевого червя система органов размножения отличается большой сложностью, сложны и связанные с размножением поведенческие реакции. Возможно, это обусловлено наземным образом жизни и необходимостью предохранять от высыхания гаметы и оплодотворенные яйца. Дождевые черви-гермафродиты (рис. 4.31). Это можно рассматривать как приспособление к относительно оседлому образу жизни. При таком образе жизни встречи дождевых червей друг с другом происходят очень редко, но если это случается, то любой представитель этого же вида может устроить дождевого червя в качестве полового партнера, поскольку оба они гермафродиты. В процессе копуляции происходит взаимооплодотворение, т. е. обмен мужскими гаметами.
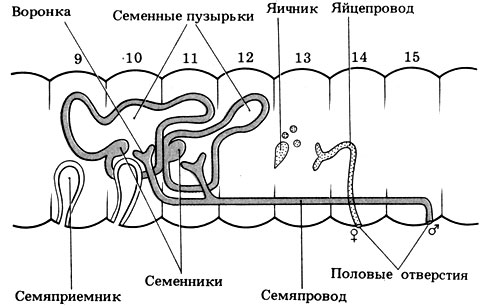
Рис. 4.31. Система органов размножения дождевого червя. Вид сбоку
Половые органы у дождевых червей сосредоточены на переднем конце тела. Расположение системы органов размножения показано на рис. 4.31. Довольно сложный процесс спаривания и последующая откладка оплодотворенных яиц в кокон происходят следующим образом.
Весной и летом теплыми влажными ночами дождевой червь высовывается из своей норы, редко оставляя ее полностью, и спаривается со своим соседом. Они соприкасаются брюшными поверхностями так, что голова одного из них оказывается направленной в сторону хвостового конца другого. При этом 9-11-й сегменты тела располагаются напротив пояска партнера. В области пояска, а также на 10-15-м и 26-м сегментах находятся длинные щетинки, которые черви вонзают друг в друга для того, чтобы обеспечить более тесный контакт во время копуляции.
Эпителий обоих червей секретирует слизистую муфту вокруг сегментов 11-31. Эти муфты разделяют сперму партнеров при копуляции; в ней возникает специальный замкнутый желобок для прохождения спермы.
В области поясков вокруг партнеров секретируется общая трубка, которая также удерживает олигохет вместе.
У обоих партнеров сперма из семенного пузырька по семяпроводу, открывающемуся наружу на 15-м сегменте, выводится наружу и по брюшному семенному желобку перемещается в направлении к заднему концу. Движение спермы по желобку обеспечивается сокращениями дугообразных мышц, расположенных в 15-32-м сегментах в слое продольной мускулатуры. Достигнув 9-го и 10-го сегментов тела партнера, сперма поступает в его семяприемники.
После того как завершается обмен спермой, партнеры расходятся (процесс занимает 3-4 ч). А через два дня начинает формироваться кокон.
Вокруг каждого червя эпителиальными железами секретируется плотная хитиновая трубка; она становится оболочкой кокона. Клетки пояска секретируют в кокон альбумин, которым впоследствии будут питаться зародыши. В результате расширения сегментов, находящихся позади кокона, он проталкивается вперед. В это время через отверстие яйцевода, находящееся на 14-м сегменте, в кокон откладываются 10-12 яиц. При движении кокона мимо 9-го и 10-го сегментов в него поступает сперма из семяприемников и происходит оплодотворение. Наконец, кокон соскальзывает с червя. Края кокона быстро смыкаются, что предотвращает высыхание его содержимого. Кокон сначала имеет желтую окраску, но по мере подсыхания темнеет.
Коконы образуются каждые 3-4 дня, до тех пор пока не будет израсходована вся сперма. Образование коконов может продолжаться в течение года без дополнительного спаривания.
Развитие у аннелид прямое, т. е. они не имеют свободноплавающих личиночных стадий. Обычно в коконе развивается только один зародыш. Молодые черви вылупляются через 2-12 нед после откладки кокона в зависимости от условий среды.
Хозяйственное значение дождевых червей
Роющая активность дождевых червей улучшает аэрацию почвы и ее дренажные свойства. Вырытые ходы облегчают рост корней в почве. Дождевые черви выносят на поверхность из более глубоких слоев частицы земли, содержащие неорганические компоненты. Таким образом они участвуют в перемешивании почвы.
Комочки, превышающие 2 мм в диаметре, черви заглатывать не могут, поэтому почва, которую они выносят на поверхность, не содержит камешков и в ней создаются хорошие условия для прорастания семян. Благодаря деятельности дождевых червей семена могут оказываться под слоем почвы, что способствует их успешному созреванию.
Дождевые черви затаскивают в норы листья, частично используя их в пищу. Остатки листьев, так же как экскременты, секреты и трупы червей, обогащают почву органическими компонентами.
Выбросы земляных червей имеют рН около 7, поэтому они предотвращают защелачивание или закисление почвы.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'