7.2. Структуры, общие для животных и растительных клеток
7.2.1. Клеточные мембраны
Клеточные мембраны играют важную роль по ряду причин. Они отделяют клеточное содержимое от внешней среды, регулируют обмен между клеткой и средой и делят клетки на отсеки, или компартменты, предназначенные для тех или иных специализированных метаболических путей. Некоторые химические реакции, в частности световые реакции фотосинтеза в хлоропластах или окислительное фосфорилирование при дыхании в митохондриях, протекают на самих мембранах. Здесь же на мембранах располагаются и рецепторные участки для распознавания внешних стимулов (гормонов или других химических веществ), поступающих из окружающей среды или из другой части самого организма. Знакомство со всеми свойствами клеточных мембран необходимо для понимания того, как функционирует клетка.
С конца прошлого века известно, что клеточные мембраны ведут себя не так, как полупроницаемые мембраны, способные пропускать лишь воду и другие малые молекулы, например молекулы газов. Клеточные мембраны обладают избирательной проницаемостью: через них медленно диффундируют глюкоза, аминокислоты, жирные кислоты, глицерол и ионы, причем сами мембраны в известной мере активно регулируют этот процесс - одни вещества пропускают, а другие нет.
Ранние работы по проницаемости мембран показали, что органические растворители, например спирт, эфир или хлороформ, проникают сквозь мембраны даже быстрее, чем вода. Это свидетельствовало о том, что в мембранах есть какая-то неполярная часть; иными словами, что мембраны содержат липиды. Позже данное предположение удалось подтвердить химическим анализом. Выяснилось, что мембраны состоят почти целиком из белков и липидов. О белках мы будем говорить ниже. Липиды в мембранах представлены фосфолипидами, гликолипидами и стеролами.
У фосфолипидов (соединений, содержащих фосфатную группу) молекулы состоят из полярной*головы и двух неполярных хвостов (рис. 5.19). Гликолипиды представляют собой продукт соединения липидов с углеводом. Подобно фосфолипидам, они состоят из полярной головы и неполярных хвостов. Стеролами называют спирты, относящиеся к классу стероидов. Наиболее распространен среди них холестерол (рис. 5.20). Его молекулы полностью неполярны, и в этом его отличие от фосфолипидов и гликолипидов.
* (Напомним, что полярные группы или молекулы несут заряд и проявляют сродство к воде (они гидрофильны), а неполярные с водой не смешиваются (они гидрофобны) (разд. 5.12).)
Если по поверхности воды распределяется тонкий слой каких-нибудь полярных липидов, например фосфолипидов, то их молекулы ориентируются таким образом, чтобы образовать один мономолекулярный слой, или монослой, как показано на рис. 7.7. Неполярные гидрофобные хвосты молекул торчат при этом из воды, а полярные гидрофильные головы лежат на ее поверхности.
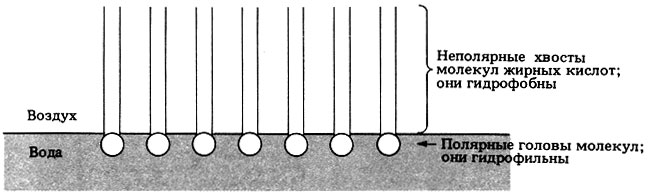
Рис. 7.7. Монослой полярных липидных молекул, например фосфолипидов, на поверхности воды
Если количество липидов больше количества, необходимого для того, чтобы покрыть поверхность воды, или если смесь липидов с водой встряхнуть, то образуются частицы, называемые мицеллами: в них гидрофобные хвосты упрятаны внутрь и тем самым защищены от контакта с водой (рис. 7.8).
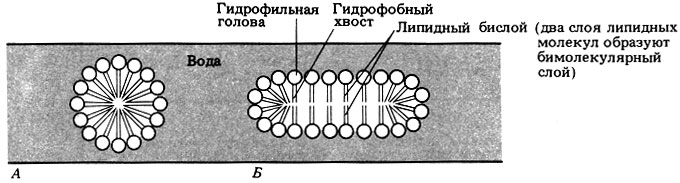
Рис. 7.8. Мицеллы, образуемые полярными липидами в воде: сферическая (А) и палочковидная (Б)
На рис. 7.8 изображена мицелла, в которой имеется двойной слой липидных молекул - так называемый липидный бислой. Подобные липидные бислои обладают многими свойствами, характерными для мембран, находящихся в живых клетках. Давсон и Даниелли (Davson, Danielli) в 1935 г. высказали предположение, что в клеточных мембранах имеется такой же липидный бислой, заключенный между двумя слоями белка. Предложенная этими авторами модель мембраны изображена на рис. 7.9. С появлением электронного микроскопа впервые открылась возможность познакомиться со строением мембран, и тогда обнаружилось, что плазматическая мембрана как животных, так и растительных клеток выглядит именно как трехслойная (триламинарная) структура. На рис. 7.10 приведена в качестве примера электронная микрофотография плазматической мембраны эритроцита.
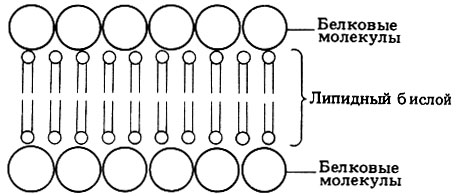
Рис. 7.9. Модель мембраны, предложенная Давсоном и Даниелли
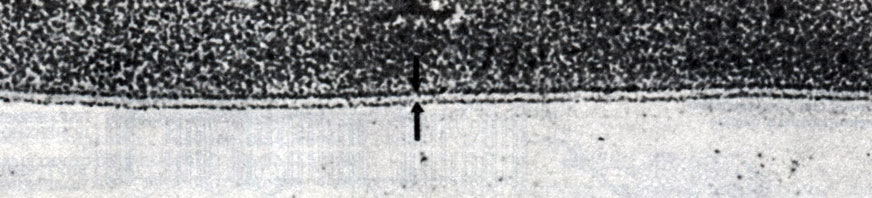
Рис. 7.10. Электронная микрофотография плазматической мембраны эритроцита; × 250000. Стрелками отмечена трехслойная структура мембраны (одна светлая полоса между двумя темными). Для фиксации использована четырехокись осмия, поглощаемая гидрофильными частями белковых и липидных молекул
В 1959 г. Робертсон (Robertson), объединив имевшиеся в то время данные, выдвинул гипотезу о строении "элементарной мембраны", в которой он постулировал структуру, общую для всех биологических мембран:
а)все мембраны имеют толщину около 7,5 нм;
б)в электронном микроскопе все они представляются трехслойными;
в)трехслойный вид мембраны есть результат имен но того расположения белков и полярных липидов, которое предусматривала модель Давсона и Даниелли (рис. 7.9), - центральный липидный бислой заключен между двумя слоями белка.
Эта гипотеза строения элементарной мембраны претерпела с тех пор изменения в связи с получением новых данных, поступавших из различных источников. Особенно плодотворным в деле изучения мембранной структуры оказался метод замораживания-скалывания, описанный в разд. П.2.5. При этом мембраны расщепляются и внутренние их поверхности становятся доступными для обозрения. Преимущество этого метода заключается в том, что мембраны благодаря мгновенному замораживанию сохраняются как бы в живом виде, тогда как химическая фиксация способна так или иначе изменить естественное расположение их компонентов. Метод замораживания - скалывания выявляет наличие в мембране частиц (главным образом белков), погруженных в липидный бислой, а иногда и пронизывающих его насквозь. В целом можно сказать, что, чем активнее в метаболическом плане данная мембрана, тем больше в ней обнаруживается таких белковых частиц; в мембранах хлоропластов (содержащих до 75% белка) частиц много (рис. 7.13), а в метаболически инертной миелиновой оболочке аксона (18% белка) их нет совсем. Неодинаково и расположение таких частиц на внутренней и наружной поверхности мембраны.
В 1972 г. Сингер и Николсон (Singer, Nicolson) предложили жидкостно-мозаичную модель мембраны, согласно которой белковые молекулы, плавающие в жидком липидном бислое, образуют в нем как бы своеобразную мозаику. Жидкостно-мозаичная модель мембраны представлена на рис. 7.11.
7.3. а) Какие структуры обозначены буквами А, В, С и D на рис. 7.11, А?
б) Какие общие компоненты структуры D здесь опущены?
В этой модели липидный бислой по-прежнему рассматривается как элементарная мембрана, но здесь он представлен как динамическая структура; белки плавают в этом липидном "море" подобно островам - иногда свободно, а иногда как бы на привязи - их удерживают микрофиламенты, проникающие в цитоплазму. Липиды также могут перемещаться, меняя свое положение.
Белки
Некоторые мембранные белки лишь частично погружены в мембрану, тогда как другие пронизывают всю ее толщу. Обычно у белков имеются гидрофобные участки, взаимодействующие с липидами, и гидрофильные участки, находящиеся на поверхности мембраны в контакте с водным содержимым клетки. В клеточных мембранах встречаются тысячи различных белков. Среди них есть чисто структурные белки и белки, выполняющие наряду со структурными также какие-либо другие, дополнительные функции. Некоторые, например, действуют как переносчики, транспортируя через мембрану те или иные вещества. Такие переносчики могут входить как составная часть в какой-нибудь активный насосный механизм (об этих механизмах мы будем говорить позднее). Предполагается, что в белковых молекулах или между соседними белковыми молекулами имеются гидрофильные каналы, или поры. Эти поры пронизывают мембрану, так что по ним сквозь мембрану могут проходить полярные молекулы, которые без таких пор пройти бы не могли - липидный компонент мембраны не пропустил бы их в клетку. На рис. 7.11, А изображена одна такая пора.
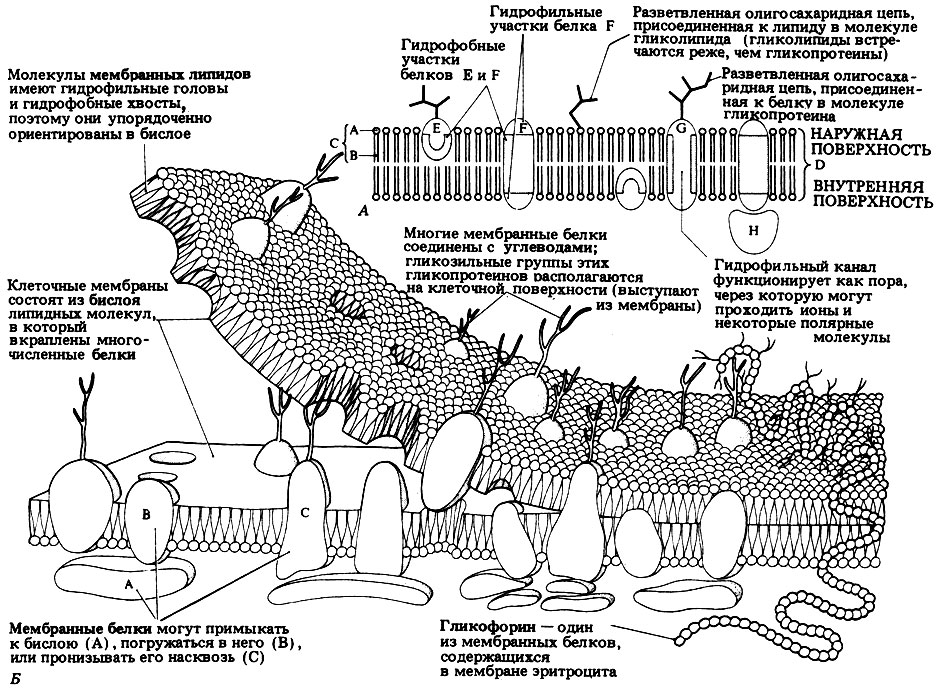
Рис. 7.11. А. Плоскостное изображение жидкостно-мозаичной модели мембраны. Гликопротеины и гликолипиды связаны только с наружными поверхностями мембраны. Б. Трехмерная модель мембраны
В мембранах содержатся ферментные белки, специфические рецепторы, переносчики электронов, преобразователи энергии, участвующие в фотосинтезе и дыхании, и т. п. Кроме того, в мембранах имеются гликопротеины. У них на свободных поверхностях находятся гликозильные группы - разветвленные олигосахаридные цепи, напоминающие антенны (рис. 7.11). Эти "антенны", состоящие из нескольких моносахаридных остатков, имеют самую разнообразную (хотя и строго определенную) конформацию, что объясняется разнообразием связей между моносахаридными остатками и существованием α- и (β-изомеров (см. гл. 5). Функция "антенн" связана с распознаванием внешних сигналов, которое важно для клеток по многим причинам. Распознающие участки двух соседних клеток могут, например, связываться друг с другом, обеспечивая сцепление клеток. Благодаря этому клетки правильно ориентируются и образуют ткани в процессе дифференцировки. С распознаванием связана и деятельность различных регуляторных систем, а также иммунный ответ, в котором гликопротеины играют роль антигенов. Распознающие участки имеются и у некоторых молекул, находящихся в растворе, благодаря чему они избирательно поглощаются клетками с комплементарными распознающими участками. О присоединении углеводных остатков к белкам (гликозилировании белков), которое и придает белкам эту способность к распознаванию и которое осуществляется в аппарате Гольджи, мы еще будем говорить подробнее (разд. 7.2.7). Сахара, таким образом, могут функционировать как информационные молекулы, т. е. в этом смысле их можно сравнить с белками и нуклеиновыми кислотами.
Липиды
Состав мембранных липидов варьирует, и это влияет на такие их свойства, как жидкое состояние и проницаемость; обычно мембранные липиды по своей консистенции напоминают оливковое масло. У ненасыщенных липидов в углеводородных хвостах молекул (разд. 5.3.1. и рис. 5.17) имеются так называемые "изломы". Эти "изломы" препятствуют слишком плотной упаковке молекул и делают структуру мембраны более рыхлой, более "жидкой". С увеличением длины углеводородных хвостов липидных молекул мембрана также становится более жидкой; в известных пределах жидкое ее состояние регулирует липид холестерол. От жидкого состояния зависит активность мембран и, в частности, легкость слияния отдельных мембран друг с другом, а также активность связанных с мембраной ферментов и транспорт белков.
Гликолипиды, подобно гликопротеинам, вносят свой вклад в формирование распознающих участков.
Общая характеристика клеточных мембран
Ниже суммированы известные нам данные, касающиеся строения биологических мембран.
1. Разные типы мембран различаются по своей толщине, но в большинстве случаев толщина мембран составляет 5-10 нм; например, толщина плазматической мембраны равна 7,5 нм.
2. Мембраны - это липопротеиновые структуры (липид + белок). К некоторым липидным и белковым молекулам на внешних поверхностях присоединены углеводные компоненты (гликозильные группы). Обычно на долю углевода в мембране приходится от 2 до 10%.
3. Липиды спонтанно образуют бислой. Это объясняется тем, что их молекулы имеют полярные головы и неполярные хвосты.
4. Мембранные белки выполняют разнообразные функции.
5. Гликозильные группы связаны с механизмом распознавания.
6. Две стороны мембраны могут отличаться одна от другой и по составу, и по свойствам.
7. Мембранные липиды и белки быстро диффундируют в латеральном направлении (в плоскости мембраны), если только они как-нибудь не закреплены или не ограничены в своем передвижении.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'