3. Продукция бактерии и термофильные бактерии
Любопытно, что первыми наблюдениями подводных оазисов, связанных с гидротермализмом, мы обязаны геофизику Лонсдейлу: по фотографиям, полученным при помощи буксируемого устройства, протянутого на расстоянии 10-30 м над дном на излияниях Галапагосского рифта и примерно на 3° с. ш. над Восточно-Тихоокеанским поднятием, он описал в 1977 году изобильные поселения крупных раковин и, потрясенный их невероятной для подобной глубины плотностью, выдвинул две первые гипотезы. Первая из них предложила чисто физическое объяснение: эмиссия гидротермального флюида, температура которого, как предполагали уже тогда, достигает 100-200 °С, вызывает образование конвективных ячеек, и частицы органического вещества из приповерхностного освещенного слоя под влиянием горизонтальных течений, возникших в результате конвекции, концентрируются в непосредственной близости от горячих источников. Вторая же гипотеза, которая тогда казалась менее правдоподобной, предполагала существование бактерий, способных быстро развиваться на некоторых соединениях, несомых гидротермальными флюидами. Уже во время первых погружений американского подводного аппарата "Алвин" на излиянии на Галапагосском рифте в 1977 году именно вторая гипотеза подтвердилась прямыми наблюдениями. Наиболее активные источники испускали опалесцирующую молочного цвета жидкость, что может указывать на наличие в ней серы в коллоидальном состоянии, серы, возникающей либо при спонтанном окислении сульфидов, либо под действием бактерий. Высокое содержание сероводорода и сульфидов разных металлов, установленное в этих флюидах, было первым указанием в пользу такой гипотезы: оно позволило предполагать, что здесь существует настоящая первичная продукция, очень локализованная и производимая автотрофными хемосинтезирующими бактериями. Если первая гипотеза сегодня не может быть окончательно отброшена, то вторая - как подтверждают многочисленные указания, верная в том, что касается участия в процессе хемосинтезирующих бактерий, - должна быть глубоко переработана в той части, которая касается использования этой продукции беспозвоночными животными. В действительности оказалось, что беспозвоночные не собирают, как думали раньше, бактерий, обитающих во внешней среде, но создают с ними весьма тесную ассоциацию: "полезные" бактерии живут внутри клеток животных, которые ими питаются!
Перед тем, как подойти вплотную к этому вопросу, следует вкратце напомнить, что же представляет собой явление автотрофного хемосинтеза. Автотрофные растения способны синтезировать необходимый им органический углерод из СО2, используя солнечную энергию, улавливаемую в ходе фотосинтеза, тогда как животные, которых называют гетеротрофными, могут лишь преобразовывать произведенные растениями органические вещества для построения своих собственных клеточных компонентов. Кроме этих двух основных типов обмена веществ, фотоавтотрофии и гетеротрофии, существует третий, совсем особый, представленный хемосинтезирующими бактериями, или просто хемотрофами. Эти бактерии, для того чтобы превратить минеральный углерод в углерод органический, используют энергию, полученную при окислении неорганических веществ. Рисунок 16 показывает место, занимаемое хемосинтезом среди разных автотрофных путей обмена веществ.
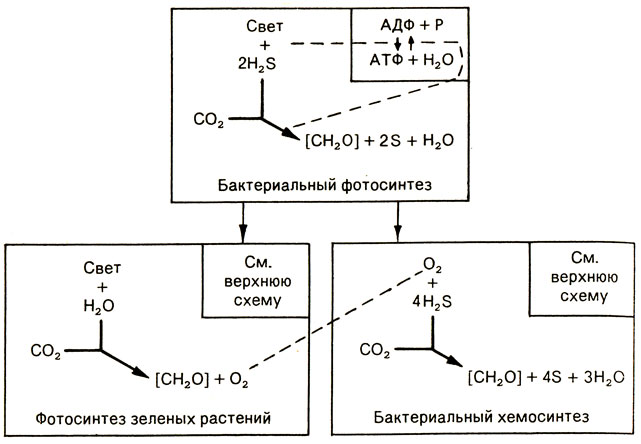
Рис. 16. Упрощенная схема положения бактериального хемосинтеза по отношению к фотосинтезу примитивных микроорганизмов и зеленых растений
Первыми фотосинтезирующими организмами были прокариоты (группа микроорганизмов, лишенных мембраны, отделяющей ядро от содержимого клетки; к ней принадлежат бактерии и синезеленые водоросли, или цианобактерии), которые жили в воздушной или жидкой среде, лишенной кислорода, и использовали свет как источник энергии для восстановления СО2, при участии сероводорода или других восстановленных соединений. 5 ходе эволюции у синезеленых, а позднее и у растений, вода заместила сероводород, а кислород соответственно серу как продукт экскреции, выбрасываемый в окружающую среду. Высвобождаемый таким образом кислород очень быстро захватил почти все биотопы суши и, как настоящий яд, вытеснил примитивные анаэробные организмы, которые вынуждены были либо искать убежища в немногих защищенных от этого "загрязнения" биотопах, либо полностью исчезнуть.
Именно на основе сероводорода, как это имело место на Земле несколько миллиардов лет назад, и функционируют хемосинтезирующие бактерии.
Первые наблюдения были сделаны на излиянии на Галапагосском рифте, в месте, где температура гидротермальных источников колеблется в пределах 15-40 °С. Прежде всего было установлено, что в пробах опалесцирующего флюида содержится очень много бактерий: 105-106 бактерий в миллилитре воды, а вблизи дна на тех же глубинах насчитывается 102-104 клеток в миллилитре воды. Повышенная изменчивость этих величин объясняется, вероятно, большей или меньшей разбавленностью воды, из которой взяты пробы. Эти цифры говорят о такой плотности бактерий, а следовательно, о таком количестве доступного для животных органического вещества (бактериальная биомасса порядка 0,1-1 мг/л), которые значительно превышают норму. Другой метод определения биомассы бактерий состоит в оценке содержания в определенном объеме морской воды аденозин-5-трифосфата (более известного под сокращенным названием АТФ), внутриклеточного компонента, играющего роль высокоэнергетического горючего. Если в нормальной придонной воде концентрация АТФ составляет 1,7 нг/л (1 нг - одна миллиардная часть грамма) при максимуме в 100 нг/л в высокопродуктивных поверхностных водах, то в пробах гидротермального флюида были получены значения 300-500 нг/л. Равным образом и другой биохимический показатель, характеризущий скорость клеточного биосинтеза, - отношение количества гуанозин-5-трифосфата к количеству АТФ (ГТФ/АТФ), измеренный в тех же пробах гидротермального флюида, оказался в четыре-пять раз выше, чем в продуктивных поверхностных водах, и в сотни раз выше, чем в обычных придонных водах. В тех же самых теплых излияниях с опалесцирующей жидкостью можно невооруженным глазом увидеть частицы, которые представляют собой бактериальные агрегаты. Один из видов рыб, свойственный излияниям Галапагосского рифта (уже упомянутый выше), часто держится прямо над источником головой вниз, будто собирая агрегаты бактерий, что являлось бы уникальным примером столь короткой пищевой цепи.
Для получения более точной оценки продукции органического вещества бактериями, как правило, измеряется количество ассимилированного СО2, меченого радиоактивным изотопом углерода 14С. С помощью манипуляторов аппарата "Алвин" удалось наполнить гидротермальным флюидом шприцы с заранее введенным в них определенным количеством радиоактивного СО2. После инкубации в течение определенного времени достаточно профильтровать воду и отделить бактерии. Величина радиоактивности, зарегистрированной в бактериальных клетках, соответствует доле меченого минерального углерода, преобразованного в органический углерод. Результаты, полученные таким методом, очень интересны: наибольшая величина продукции, полученная при температуре около 3-4 °С, то есть после сильного разбавления гидротермального флюида окружающей морской водой, составляет 3,6-4,8 мг Углерода на 1 м3 воды в сутки, что примерно соответствует той величине, какую можно получить в глубоком аноксическом бассейне между насыщенным кислородом поверхностным слоем и глубинным слоем, например во впадине Карьяко в Карибском море. Эти величины не покажутся очень высокими, если сравнить их с 11,7 мг ассимилированного углерода на 1 м3 в сутки, установленными для глубоководных слоев Черного моря, то есть для среды, лишенной кислорода в результате естественных причин. Исходя из повышенной концентрации сероводорода в гидротермальном флюиде, пытались также доказать наличие бактерий, способных окислять сульфиды. Среди сотен штаммов бактерий, изолированных и выделенных в культуры, многие оказались способны окислять сероводород: одни из них - это облигатные хемотрофы, их метаболическая активность приводит к выделению в среду кислых веществ; другие - факультативные хемотрофы, они выделяют основные вещества (политионаты). Удалось определить некоторые штаммы, переносящие повышенное содержание сероводорода при термическом оптимуме 25-35 °С, но при давлении 250 бар (давление, соответствующее глубине излияния на Галапагосском рифте) они сохраняют только 75% своей активности на поверхности (например, бактерии, принадлежащие к роду Thiomicrospira).
Общий вывод из исследований, проведенных на излияниях Галапагосского рифта, ясен: в физико-химических условиях, свойственных этому местонахождению, ассимиляция СО2 (мера хемосинтетической активности) в гидротермальном флюиде в ходе перемешивания его с окружающей морской водой низка по сравнению с биомассой бактерий, обнаруженных в этом флюиде. Следовательно, эта биомасса бактерий, по всей вероятности, создается в подповерхностной сети трещин и разломов, во всяком случае до выхода флюида в морскую воду. Микробиологи даже считают, что метаболическая активность бактерий, присутствующих в гидротермальном флюиде, в общем много меньше той, которая могла бы удовлетворить нужды процветающих сообществ животных.
Не следует ли вернуться к первой гипотезе, согласно которой конвективные ячейки, порожденные гидротермальной эмиссией, способствуют тому, что как раз на уровне излияний концентрируется достаточное количество взвешенного органического вещества, приносимого с поверхности моря? Нет, не следует, ибо простые расчеты показывают, что количество органического вещества, которое подобным образом может постоянно поступать сюда, намного ниже потребностей плотных сообществ животных. В действительности бактериальная продукция, создающаяся в основном внутри гидротермальной сети, под поверхностью базальтов, вероятно, достаточна, чтобы объяснить большую плотность поселений некоторых беспозвоночных-фильтраторов, способных улавливать взвешенные частицы, и в частности агрегаты бактерий. Это объясняет также наличие мелких планктонных животных над излияниями.
Опять же в пробах, взятых главным образом на Галапагосском рифте, были определены разные типы хемосинтезирующих бактерий. Различают шесть крупных морфологических категорий:
- Одноклеточные бактерии округлой формы, часто заключенные в плотные железисто-марганцевые отложения.
- Бактерии, имеющие очень длинную ножку, типа Hyphomonas-Hyphomicrobium.
- Формы, напоминающие цианобактерии, морфолочески очень близкие к Calothrix. - Формы, содержащие сильно развитые цитоплазмаческие мембранные системы, определенные как метилотрофные бактерии.
- Нитчатые бактерии, напоминающие роды fhiotrix и Leucothrix, которые составляют большинство проанализированных бактерий.
- Нитчатые неприкрепленные бактерии, напоминающие Beggiatoa.
На излияниях черных курильщиков, и в частности на местонахождении 21° с. ш. на Восточно-Тихоокеанском поднятии, было проведено меньше исследований. Здесь были получены более оригинальные, но порой спорные результаты. Во-первых, удалось показать существование термофильных бактерий, то есть таких бактерий, у которых оптимальная метаболическая активность наблюдается при температуре выше 50 °С. Для некоторых проб бактерий был установлен температурный оптимум 105 °С; эти экстремально термофильные бактерии при температурах ниже 70 °С становятся неактивными. Метаболическая активность этих бактерий чрезвычайно интересна: в культуральных средах, содержащих минералы (S2O3, Мn и Fe), ПРИ температуре 100 °С и давлении 1 атмосфера они способны синтезировать метан (СН4), окись углерода (СO) и водород, а также небольшое количество закиси азота (N2O); они могут также окислять метан. Если до этого открытия геологи считали, что метан, водород и окись углерода, содержащиеся в гидротермальных флюидах, возникают либо при дегазации мантии Земли, либо в результате химических реакций между базальтом и морской водой, то это открытие объясняет явление с биологической точки зрения. Эти термофильные бактерии быстро размножаются: штамм, принадлежащий к роду Methanococcus, обнаруженный в отложениях минеральных частиц в основании черного курильщика, имеет оптимум роста при 86 °С и время удвоения 26 минут.
Но вернемся к бактериям, выделенным из гидротермального флюида черного курильщика, взятого при температуре 350 °С: в июне 1983 года в авторитетном научном журнале "Нейчер" ("Природа") появилась статья, всего на три страницы, озаглавленная "Рост бактерий из черных курильщиков при температуре как минимум 250 °С"! Эта короткая статья знакомит с опытами, проведенными с пробой воды, взятой осенью 1979 года на Восточно-Тихоокеанском поднятии на 21° с. ш. из султана черноватой жидкости температурой 306 °С. Бактериальные культуры были выращены в камере, позволяющей воссоздать температуру и давление, свойственные этому местонахождению (то есть температуру 350 °С и давление 265 атмосфер), на среде, в состав которой входили различные минеральные соли. При подсчете бактерий в эпифлуоресцентном микроскопе (процедура состоит в стимуляции флуоресценции красителем DAPI*, который избирательно фиксируется на молекулах ДНК хромосомного вещества бактерий) были установлены следующие скорости удвоения бактерий: 8 часов при температуре 150 °С; 1,5 часа при температуре 200 °С и всего лишь 40 минут при температуре 250 °С. При последнем значении температуры масса белка удваивается с той же скоростью, что и число бактерий. При температуре 300 °С после некоторого возрастания количества клеток наблюдается довольно быстрое его уменьшение, что может быть объяснено разрушением клеток во время необходимой для взятия проб декомпрессии или даже во время самого опыта, поскольку, в частности, при высоких давлении и температуре в морской воде понижается показатель кислотности. На некоторых снимках, сделанных в электронном микроскопе, видны организмы, выросшие в культуре при температуре 250 °С и давлении 265 атмосфер.
* (4,6-диамидино-2-фенилиндол. - Прим. ред.)
Чтобы лучше понять значение этой публикации и тот отпор, который ей дали микробиологи, необходимо сделать некоторые пояснения общего характера. Самая высокая экспериментально измеренная температура, при которой возможны жизнь и рост микроорганизмов, в начале века составляла 55 °С, в 1970 году - 95 °С и в 1982-м - 105 °С. Последний результат был получен на бактериях, взятых из геотермальной скважины на морской сольфатаре на малой глубине, а также, как говорилось выше, на некоторых штаммах, изолированных из проб черноватого гидротермального флюида. Только микроорганизмы - имеются в виду прокариоты - способны жить при температурах такого порядка. В начале 1970-х годов один микробиолог, специалист по термофильным бактериям, на основе изучения микрофлоры гейзеров Йеллоустонского Национального парка выдвинул идею о том, что наличие воды в жидком состоянии - гораздо более важное условие для существования жизни, чем максимальная температура. Все бактерии, способные переносить температуры выше 80-85 °С, принадлежат к группе архебактерий, которые, как указывает их название, считаются наиболее близкими к предковым формам: много среди них метаногенных бактерий, способных синтезировать метан (роды Thermoplasrna, Sulfolobus, Methanococcus). Недавно описанный вид из пробы черноватого гидротермального флюида, Methanococcus jannaschii, отличается от других известных архебактерий оригинальной биохимической структурой клеточной мембраны: у архебактерий мембранные липиды состоят из изопреноидных углеводородов и алкилглицериновых эфиров, а у вновь открытого вида мембранные липиды на 95% состоят из макроциклического диэфира глицерина - молекулы, доселе не встречавшейся в живом мире.
Палеонтологам давно знакомы ископаемые структуры, являющиеся результатом жизнедеятельности морских микроорганизмов, - строматолиты, наиболее древние из ныне известных свидетельств жизни. Некоторые строматолиты имеют возраст около 3,5 млрд. лет и отложения, в которых они находятся, иногда носят следы гидротермальной деятельности. Эти факты следует рассматривать в общем контексте эволюции нашей планеты: миллиард лет тому назад примитивный океан был совсем лишен кислорода, и его температура, по мнению некоторых специалистов, должна была составлять 50-70 °С. В более отдаленные эпохи температура океана, возможно, была еще выше и достигала 150 °С, оставаться жидкой при такой температуре воде позволяло высокое давление земной атмосферы, связанное с очень большим содержанием СО2. Задолго до образования континентов этот архейский океан, похоже, был совсем неглубоким и являлся ареной интенсивных гидротермальных проявлений.
Из этого следует, что микроорганизмы, которые жили на Земле первые два миллиарда лет после возникновения жизни, были термофильными формами, обитавшими в аноксической среде и использовавшими для своего развития минеральные элементы, происходящие из базальтовой магмы. В той особенной среде, которая представляет собой гидротермальную подповерхностную сеть, до наших дней сохраняются физико-химические условия если не идентичные, то очень близкие к условиям архейского океана. В гидротермальных выходах, относящихся к этой сети, до сих пор находят архебактерий, и можно утверждать, что изучение этой среды представляет особый интерес для познания условий развития первых сообществ микроорганизмов.
Не удивительно поэтому, что сообщение об открытии бактерий, способных размножаться при температуре 250 °С и давлении 265 атмосфер, стимулировало исследование этих проблем и повлекло за собой серию работ, призванных проверить полученные результаты. В самом деле, если бы они подтвердились, была бы поставлена под сомнение распространенная теория происхождения жизни на Земле, согласно которой первые органические молекулы образовались фотохимическим способом из газов, содержащихся в атмосфере, аминокислоты появились в результате конденсации углеводов и формамида, и все это привело к появлению настоящего органического "супа", плавающего на поверхности вод. В этом примитивном "супе" якобы зародились первые молекулы, способные к самовоспроизведению (нуклеиновые кислоты), затем первые прокариотные организмы. Эта теория не учитывает недавние открытия подводных гидротерм, но является в настоящее время официально принятой. Отсюда становится понятным то значение, которое биологи придают этим новым результатам. С момента выхода в 1983 году упомянутой статьи в США развернулась полемика.
Многочисленные последующие работы опровергли реальность полученного результата. Так, один биохимик показал, что максимальная температура, которую могут переносить живые существа, - это такая температура, при которой наступает естественный гидролиз их биохимических компонентов (гидролиз - химическое разложение молекулы путем фиксации воды), а температура гидролиза, по мнению исследователя, ниже 250 °С. Поэтому он приходит к благоразумному заключению: если ультратермофильные бактерии действительно существуют и их метаболические реакции происходят в водной среде, то они не могут жить при температуре 250 °С, если состоят из биологических молекул, таких, как белки и нуклеиновые кислоты. Можно критиковать и эту работу, основанную на опытах, проведенных при давлении 39,2 атмосферы (давление водяного пара при температуре 250 °С) и на изолированных молекулах, не заключенных в биологическую мембрану. Другие авторы показали, что штамм термофильных архебактерий (Pyrodictium occultum), способных жить при температуре 110 °С при атмосферном давлении в широком диапазоне солености, разрушается гидролизом за 6 часов при температуре 250 °С под давлением 260 атмосфер.
Какой же вывод можно сделать из этих работ, проведенных с таким тщанием? Для многочисленных микробиологов этих доказательств достаточно: микроорганизмов, способных жить при температуре 250 °С и давлении 265 атмосфер, не существует, а результаты, опубликованные в 1983 году, должны быть отнесены к экспериментальным артефактам. Для других вопрос остается открытым, здесь необходимы новые исследования.
Однако доказано, что в гидротермальных флюидах с высокой температурой существуют бактерии, у которых температурный оптимум жизнедеятельности близок к 110 °С. Из недавней работы Деминга, одного из двух авторов первоначального открытия ультратермофильных бактерий, доложенной на совещании в Океанографическом институте (Париж, ноябрь 1985 года), следует, что те же самые бактерии размножаются при температуре 120 °С под давлением 200 атмосфер, при культивировании их на новой твердой среде, которая остается стабильной при этой температуре. Этот результат не противоречит наблюдениям над бактериями, полученными из морских прибрежных сольфатар в Тирренском море. Откуда появляются бактерии, обнаруживаемые в гидротермальных флюидах? Если они действительно происходят, как на это указывают все имеющиеся данные, из подповерхностной гидротермальной сети, значит, в той или иной форме эти существа живут при очень высоких температурах.
Одна из проблем, связанных с этим спором, чисто техническая: каким образом из хорошо уравновешенного подводного аппарата взять металлическим батометром, заканчивающимся управляемой трубкой для забора воды, пробу гидротермального флюида, не разбавленного морской водой? Проблема эта совсем не простая, до настоящего времени она не нашла решения. Прибавим, что число проб черноватого флюида, изученных микробиологами, вряд ли превышает десяток, и тогда станет ясно, что окончательное заключение формулировать рановато. И хотя для выводов время еще не пришло, тем не менее с самой общей точки зрения тот факт, что до наших дней сохранились первоначальные физико-химические условия, существовавшие при появлении жизни на нашей планете три с половиной миллиарда лет назад, заслуживает не мимолетного внимания.
Бактерии, о которых мы вели речь, живут или свободно, или прикрепляясь к разным субстратам, или взвешенными в морской воде. В подводных оазисах есть и другие бактерии, более или менее тесно связанные с различными гидротермальными беспозвоночными. Эти ассоциации, составляющие, пожалуй, наиболее яркую особенность гидротермальных сообществ, будут предметом обсуждения в двух последующих главах.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'