17.4.8. Снабжение энергией
Обычно первоисточником энергии для мышечного сокращения служит гликоген, а иногда используются жирные кислоты. В процессе расщепления этих субстратов вырабатывается АТФ, гидролиз которого доставляет энергию непосредственно для самого сокращения:
В покоящейся мышце количество АТФ невелико, оно может обеспечить энергией лишь около восьми одиночных сокращений. Такой уровень АТФ может поддерживаться при обычном аэробном метаболизме (дыхании). В процессе работы мышц АТФ быстро расходуется, и его уровень должен быстро восстанавливаться уже за счет других процессов.
В восстановлении уровня АТФ участвует содержащийся в мышцах креатинфосфат (КрФ). Образующийся при мышечном сокращении АДФ вновь фосфорилируется в АТФ за счет креатинфосфата:
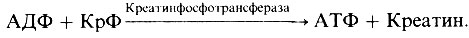
Этот процесс непрерывно пополняет запасы АТФ, которые могут использоваться непосредственно при сокращении. Но в конце концов возникает необходимость и в пополнении резервов креатинфосфата, и оно происходит за счет окисления жирных кислот или гликогена. Образующийся при этом АТФ используется для ресинтеза креатинфосфата из креатина (Кр):
При очень интенсивной работе мышц поступающего кислорода вскоре оказывается недостаточно для поддержания на должном уровне окислительного форфорилирования (разд. 11.3.6). В этих условиях пировиноградная кислота - конечный продукт гликолиза - при участии ионов Н+ превращается в молочную кислоту.
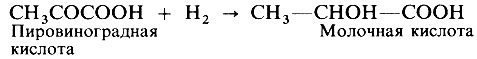
Так происходит из-за нехватки кислорода, связывающего ионы Н+ . В этом случае говорят, что работа мышцы создает кислородную задолженность.
Образование молочной кислоты - процесс с относительно малым энергетическим выходом: высвобождаемая энергия составляет лишь 7% от энергии, получаемой при полном окислении глюкозы. Молочная кислота токсична и рано или поздно должна быть выведена из организма или переработана. Так и происходит при уменьшении или прекращении мышечной работы. В этот период кислорода становится достаточно для окисления молочной кислоты и частичного превращения ее в гликоген. Такой процесс обычно происходит в печени; здесь 1/5 часть молочной кислоты полностью окисляется до углекислоты и воды; высвобождаемая при этом энергия расходуется на превращение остальной молочной кислоты в глюкозу. Часть глюкозы возвращается в мышцы, и там из нее в конце концов образуется гликоген, а глюкоза, оставшаяся в печени, превращается в гликоген и запасается в ней. Время полной переработки молочной кислоты - это именно то время, которое необходимо для ликвидации кислородной задолженности после энергичной работы мышц (рис. 17.25).
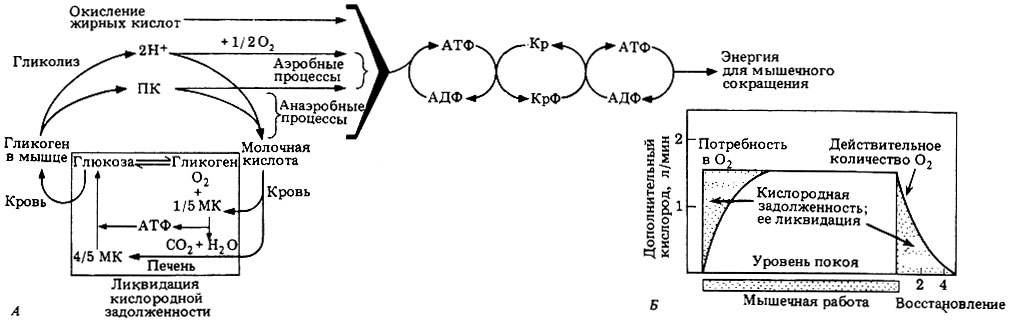
Рис. 17.25. А. Отношения между АТФ, креатинфосфатом и процессом дыхания при сокращении мышцы. МК - молочная кислота; ПК - пировиноградная кислота; Кр - креатин; КрФ - креатинфосфат. Б. Потребность в кислороде при физической работе. Показано соотношение между потреблением кислорода и созданием кислородной задолженности
17.5. Иннервация скелетной мышцы
К каждой мышце подходит много двигательных аксонов, которые ветвятся и иннервируют группы мышечных волокон. Каждая такая группа вместе с иннервирующим ее аксоном называется двигательной единицей. При надлежащей стимуляции все мышечные волокна в ней сокращаются одновременно. Число этих волокон в двигательной единице варьирует и зависит от того, насколько сложным должно быть управление данной мышцей. Например, в глазодвигательной мышце это число составляет около 10, а в бицепсе - более 1000. Чем меньше число волокон в двигательной единице, тем более тонкий контроль осуществляется со стороны нервной системы.
В месте контакта двигательного аксона с мышечным волокном формируется нервно-мышечное соединение, или двигательная концевая пластинка. Здесь аксон не имеет миелиновой оболочки и его ветвящиеся окончания погружены в разветвленные бороздки концевой пластинки.
Сигнал для сокращения поперечнополосатых мышц поступает из центральной нервной системы (рис. 17.26). Импульсы по двигательному нерву передаются на двигательную концевую пластинку. Здесь в ответ на нервный сигнал в синаптическую щель, отделяющую моторный аксон от сарколеммы мышечного волокна, секретируется ацетилхолин. События, происходящие в концевой пластинке и в межнейронных синапсах (разд. 16.1.2), удивительно сходны. Ацетилхолин диффундирует к сарколемме и на короткое время повышает ее проницаемость для ионов, особенно для Na+ и К+; в результате возникает потенциал концевой пластинки, в свою очередь генерирующий потенциал действия; последний быстро распространяется по всей длине и в глубь мышечного волокна по Т-системе, вызывая сократительную реакцию по принципу "всё или ничего".
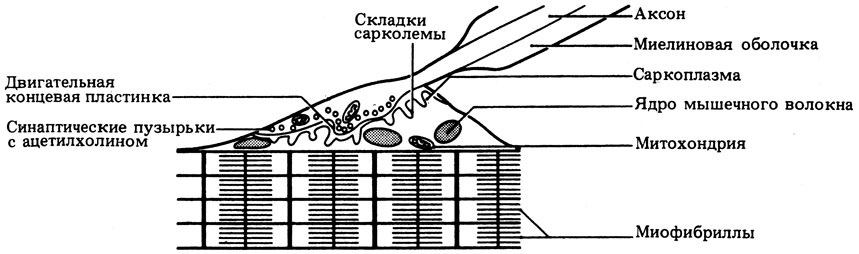
Рис. 17.26. Двигательная концевая пластинка в месте контакта нерва и мышечного волокна
В области сарколеммы в большом количестве содержится ацетилхолинэстераза, которая быстро расщепляет ацетилхолин на холин и уксусную кислоту и таким образом предотвращает избыточную стимуляцию мышечного волокна. Передав возбуждение мышечному волокну, концевая пластинка быстро возвращается в состояние покоя.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'