22.8.5. Роль генов в развитии
Связь между определенными участками молекулы ДНК и морфологическим развитием усиленно изучалась на организмах, обладающих гигантскими (политенными) хромосомами. Такие хромосомы имеются, например, в клетках слюнных желез у личинок многих двукрылых, в том числе дрозофилы. Как эти политенные хромосомы достигают таких размеров, объясняется в разд. 23.5.1. Гигантские хромосомы относительно легко изучать с помощью светового микроскопа, и окраска по Фёльгену выявляет в них специфическую для каждого участка поперечную исчерченность. В процессе развития личинки дрозофилы проходят несколько стадий ("возрастов"), разделенных линьками-периодами интенсивной активности эпидермиса, за которой следует сбрасывание старой кутикулы. Две последние линьки сопровождаются особенно резкими изменениями; одна из них ведет к образованию куколки, а другая - к выходу взрослой особи (имаго). Для этих стадий характерны интенсивный метаболизм и процессы дифференцировки. Ход развития регулируется гормонами (см. разд. 21.4). На определенных стадиях некоторые из поперечных полос на хромосомах увеличиваются и образуют структуры, известные под названием пуфов или колец Бальбиани (по имени ученого, впервые обнаружившего их в 1890 г.). Специфическое окрашивание, выявляющее РНК, и радиоавтографические исследования с использованием меченых предшественников РНК показали, что в пуфах происходит синтез РНК. Было также установлено, что величина пуфов находится в прямой зависимости от скорости синтеза РНК. Полагают, что пуфы образуются в результате раскручивания молекул ДНК, разделения комплементарных цепей и синтеза мРНК в процессе транскрипции.
В процессе развития особи наблюдается определенная последовательность образования пуфов, регулируемая гормоном линьки - экдизоном. На разных личиночных стадиях и на стадии куколки пуфы возникают в различных участках хромосом; это позволяет предположить, что они соответствуют структурным генам. Доказательства того, что это участки генетической активности, представил Беерман, изучавший личинок двух видов комаров, принадлежащих к роду Chironomus. Некоторые клетки слюнных желез у личинок одного вида зернистые, а у другого вида - нет. Генетические карты, построенные по данным скрещиваний между этими двумя видами, показали, что аллели, определяющие эти признаки, расположены вблизи центромеры одной из хромосом, причем аллель зернистости доминирует. Исследование хромосом этих видов показало, что у вида, обладающего зернистыми клетками, пуфы образуются только в том участке, где локализован этот ген. Кроме того, у личинок, гетерозиготных по признаку зернистости, пуф в этом локусе возникает только в хромосоме, несущей доминантный аллель.
Дальнейшим подтверждением связи между хромосомными пуфами и синтезом мРНК служат результаты введения организмам, обладающим гигантскими хромосомами, актиномицина D. Актиномицин D подавляет транскрипцию, препятствуя синтезу мРНК, и у особей, которым вводили это вещество, пуфы не образуются (рис. 22.35).
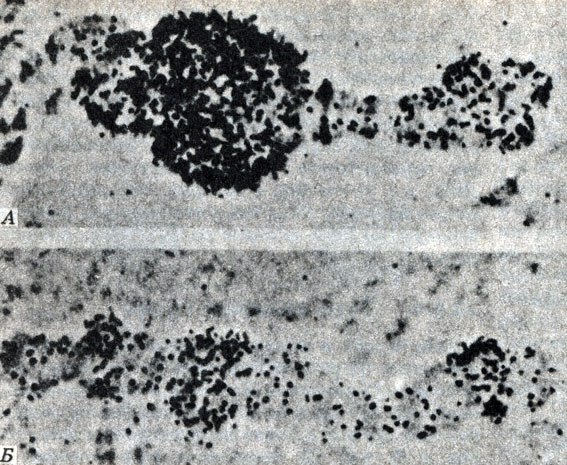
Рис. 22.35. Радиоавтографы четвертой хромосомы личинки Chironomus tentans, демонстрирующие связь между пуфингом и синтезом мРНК. Черные точки-радиоактивный уридин, включенный в процессе синтеза мРНК. А. Как показывает радиоавтограф, синтез РНК сосредоточен в области пуфа. Б. После добавления небольших количеств актиномицина D. подавляющего синтез мРНК, как пуфинг. так и включение уридина выражены слабо
На рост и развитие растений и животных оказывают также влияние гормоны. Во многих случаях их действие осуществляется на уровне транскрипции, т.е. синтеза мРНК. Конкретные механизмы воздействия различных гормонов на транскрипцию и на синтез белка весьма разнообразны. Некоторые гормоны присоединяются к рецепторным участкам клеточной мембраны. После связывания гормона рецептором, который связан с лежащим под ним ферментом аденилатциклазой, образуется циклический АМФ, который переходит в цитоплазму и действует как "второй посредник", индуцирующий транскрипцию. Подробнее об этом механизме говорится в разд. 16.6.1.
В этой главе мы попытались описать некоторые процессы, связанные с непрерывностью жизни. Живым системам, очевидно, необходима как генетическая стабильность в малых временных масштабах, так и генетическая пластичность в долговременном плане. Стабильность обеспечивают механизмы митоза, тогда как мейоз создает огромную генетическую изменчивость, позволяющую организмам приспосабливаться к меняющейся среде.
В следующей главе будут описаны особенности наследования признаков, правила, которым оно подчиняется, и пути распространения генетических изменений в популяциях.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'