23.4. Генетические карты
Данные о частотах рекомбинаций важны прежде всего потому, что дают генетикам возможность составлять карты относительного расположения генов в хромосомах. Хромосомные карты строятся путем прямого перевода частоты рекомбинаций между генами в предполагаемые расстояния на хромосоме. Если частота рекомбинации между генами А и В равна 4%, то это означает, что они расположены в одной и той же хромосоме на расстоянии 4 морганид друг от друга (морганида - единица расстояния на генетической карте); если частота рекомбинации между генами А и С равна 9%, то они разделены расстоянием 9 морганид. Однако эти данные еще не позволяют сказать, в каком порядке расположены гены А, В и С (рис. 23.12).
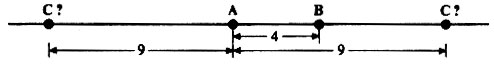
Рис. 23.12. Определение возможного положения генов А, В и С в хромосоме, основанное на данных, изложенных в тексте
На практике обычно определяют частоту рекомбинации по меньшей мере для трех генов одновременно; этот метод, называемый триангуляцией, позволяет определить не только расстояния между генами, но и их последовательность. Рассмотрим, например, частоты рекомбинации, установленные в результате ряда экспериментальных скрещиваний при участии четырех генов:
Р - Q = 24%
R - Р = 14%
R - S = 8%
S - Р = 6%
Для того чтобы установить последовательность генов и расстояния между ними, вычерчивают линию, изображающую хромосому, и производят следующие действия.
1. В середину хромосомы помещают гены с наименьшей частотой рекомбинации, т. е. S - Р = = 6% (рис. 23.13,I).
2. Выбирают следующую по величине частоту рекомбинации, т.е. R - S = 8%, и указывают два возможных положения R в хромосоме по отношению к S (рис. 23.13,II).
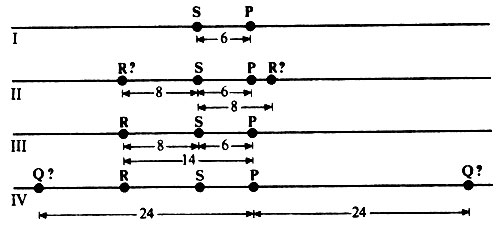
Рис. 23.13. Установление положений генов Р, Q, R и S в хромосоме с помощью метода триангуляции
3. Проделывают то же самое со следующей частотой рекомбинации, т.е. R - Р = 14%. При этом выясняется, что R не может находиться вправо от Р (рис. 23.13,III).
4. Проделывают то же самое для Р - Q = 24% (рис. 23.13,IV). Положение Q не может быть установлено без дополнительной информации. Если, например, окажется, что частота рекомбинации Q - Я =10%, то это подтвердит расположение гена Q в левом конце хромосомы.
При построении хромосомных карт могут возникнуть затруднения, создаваемые двойным кроссинговером; особенно это касается тех случаев, когда изучаемые гены разделены большими расстояниями, так как число выявляемых рекомбинантов бывает при этом меньше фактического числа перекрестов. Если, например, произойдет кроссинговер в двух местах - между А и В и между В и С (рис. 23.14), то А и С внешне проявят себя как сцепленные, но хромосома будет нести теперь рецессивный аллель b.
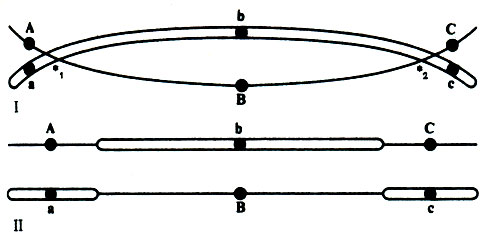
Рис. 23.14. Пора гомологичных хромаmид, одна из которых несет доминантные аллели А, В и С, а другая - рецессивные аллели a, b и с. Кроссинговер происходит в двух точках -* 1 и *2. II. Результат разделения хроматид: последовательности аллелей в них иные, хотя последовательность генных локусов и расстояния между ними остаются прежними
23.9. У кукурузы гены окрашенного семени и гладкого эндосперма доминируют над генами неокрашенного семени и сморщенного эндосперма. Линию, гомозиготную по обоим доминантным признакам, скрещивали с линией, гомозиготной по рецессивным признакам; при анализирующем скрещивании F2 с растениями F1 были получены следующие результаты:
окрашенные семена, гладкий эндосперм 380 неокрашенные семена, сморщенный эндосперм 390 окрашенные семена, сморщенный эндосперм 14 неокрашенные семена, гладкий эндосперм 10
Вычислите расстояние в морганидах между генами окраски семян и характера эндосперма.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'