Первичный метаболизм
Каким же должен был быть метаболизм первых живых организмов на Земле? Эти организмы появились "в отсутствие воздуха" в том смысле, как мы представляем воздух сегодня. Окружавшая их атмосфера содержала метан и аммиак, а не кислород и азот. Поэтому они должны были получать энергию за счет анаэробного (т. е. безкислородного) разложения молекул пищи. Как ни странно, способность к анаэробному образованию энергии сохранилась во всех живых организмах до настоящего времени, несмотря на то что к арсеналу поддерживающих жизнь обменных процессов добавились новые, более эффективные пути получения энергии.
Этот древнейший механизм обмена включает набор ферментов для расщепления молекул глюкозы на две молекулы пировиноградной кислоты. Каждый живой организм Земли - бактерия, растение и животное - имеет этот набор ферментов. Это поразительное свидетельство единства жизни на Земле.
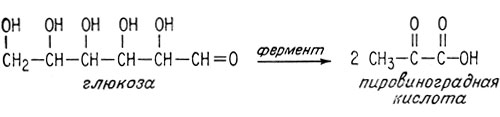
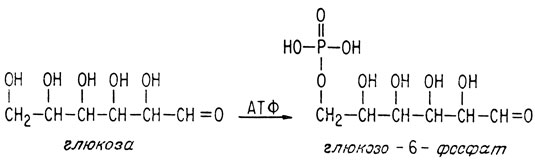
Анаэробное превращение глюкозы в пировиноградную кислоту требует нескольких стадий, каждую из которых катализирует специальный фермент. На первой стадии глюкоза превращается в глюкозо-6-фосфат, получая этот фосфатный остаток от очень важной "богатой энергией" молекулы аденозитрифосфата (АТФ) (см. далее).
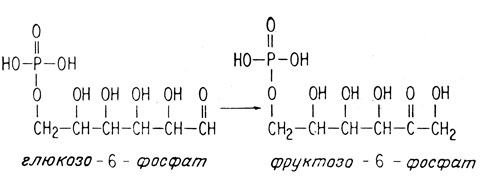
Затем глюкозо-6-фосфат изомеризуется, образуя фруктозо-6-фосфат.
Вслед за этим происходит второе фосфорилирование, при котором образуется фруктозо-1,6-дифосфат.
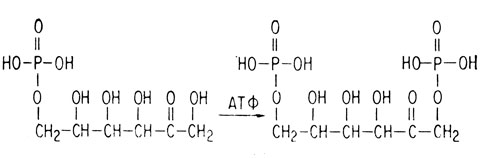
Ни одна из этих реакций фосфорилирования не поставляет организму энергию. В самом деле, при каждом фосфорилировании используется "богатая энергией" молекула аденозинтрифосфата (АТФ).
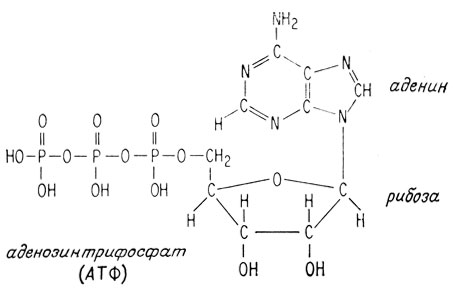
АТФ называют "богатой энергией" молекулой потому, что его концевая фосфат-фосфатная ангидридная связь является чрезвычайно реакционноспособной. Она дает энергию или является движущей силой для многих реакций обмена. Эта молекула - универсальное хранилище легкодоступной энергии всех живых организмов.
АТФ "расходует" свою богатую энергией связь, гидролизуясь до аденозиндифосфата (АДФ) и фосфорной кислоты. Суммарно эту реакцию можно записать как АТФ→АДФ+Ф. Однако при образовании фосфатов Сахаров эта энергия не расходуется на гидролиз, а используется для образования фосфатов за счет реакции "переноса фосфорильной группы" на гидроксильные группы сахара, а не воды.
В результате оставшегося от древнего механизма обмена - процесса превращения фруктозо-1,6-дифосфата в эфир пировиноградной кислоты - четыре молекулы АДФ превращаются в четыре молекулы АТФ. Таким образом, каждый раз, когда в организме происходит анаэробное разложение молекулы глюкозы до эфира пировиноградной кислоты, его запас энергии увеличивается благодаря образованию двух новых молекул АТФ.
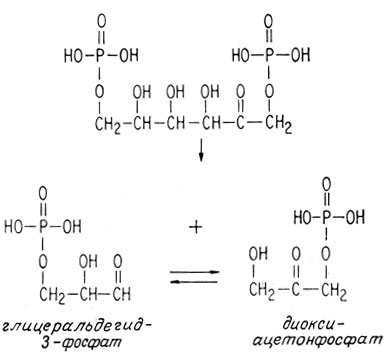
Сначала расщепляется фруктозо-1,6-дифосфат, образуя два взаимопревращаемых фрагмента, состоящих из трех атомов углерода.
Эти новые две молекулы превращаются друг в друга посредством изомеризации такого же типа, как между глюкозо-6-фосфа-том и фруктозо-6-фосфатом. Карбонильная группа перемещается на один атом с образованием промежуточного ненасыщенного диола.
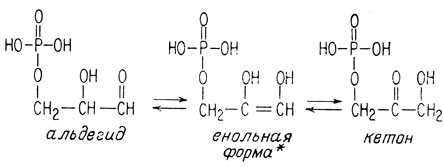
* (Процесс енолизации рассмотрен в приложении.)
В дальнейшем процессе обмена принимает участие альдегид, в данном случае глицеральдегид-3-фосфат. Следующей стадией опять является фосфорилирование, однако другого типа, чем две рассмотренные ранее реакции фосфорилирования. Эта реакция окислительного фосфорилирования и в ней не используется АТФ. Вместо этого в ней участвует обычная фосфорная кислота и чрезвычайно важный окисляющий агент, повсеместно встречающийся в природе, НАД+, или никотинамидадениндинуклеотид. Эта молекула содержит следующие звенья в указанной последовательности:
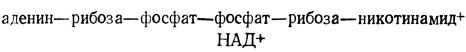
Новым звеном в этой структуре является никотинамид, представляющий собой наиболее важную часть, поскольку именно он является окисляющим участком. Можно представить эту молекулу как никотинамид, связанный с аденозиндифосфатом и рибозой (которые в приведенной ниже формуле обозначены через R).
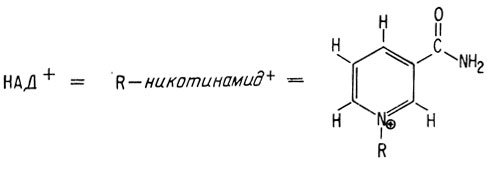
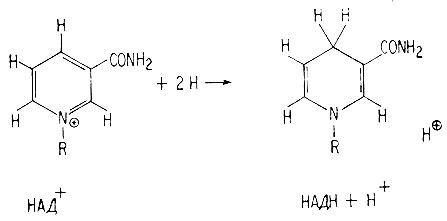
НАД+ окисляет молекулу, отщепляя от нее два атома водорода. В результате НАД+ превращается в НАДН+Н+.
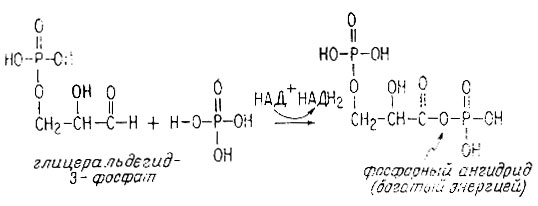
Когда глицеральдегид-3-фосфат подвергается окислительному фосфорилированию посредством НАД+ и фосфорной кислоты, он превращается в фосфорный ангидрид 3-фосфоглицериновой кислоты (альдегидная группа при этом окисляется до карбоксильной).
Значение этой стадии для организма состоит в том, что фосфорноангидридная группа, так же как и АТФ, является "богатой энергией" и может быть использована для превращения АДФ в АТФ для создания запаса энергии. Это происходит следующим образом:
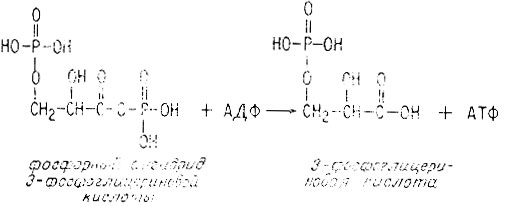
Поскольку из каждой молекулы глюкозы образуются две молекулы фосфоглицериновой кислоты, при этом получаются также две молекулы АТФ. Таким образом до этих пор в метаболизме участвовали две молекулы АТФ и были получены две молекулы АТФ в итоге изменение запаса энергии равно нулю. Однако следующие две реакции ведут к превращению 3-фосфата в богатый энергией фосфат, дальнейшее превращение которого приводит к образованию еще двух молекул АТФ, в результате чего общий запас энергии увеличивается на две молекулы АТФ на каждую молекулу глюкозы. Этот запас энергии сохраняется в высокореакционноспособных фосфорноангидридных связях АТФ.
В ходе первой из этих двух реакций фосфатная группа мигрирует от третьего ко второму гидроксилу.
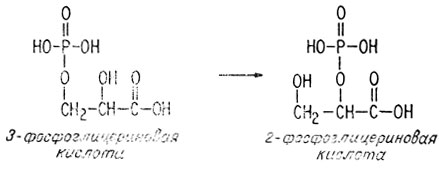
В ходе второй реакции рассматриваемая молекула теряет воду, образуя двойную связь между вторым и третьим атомами углерода.
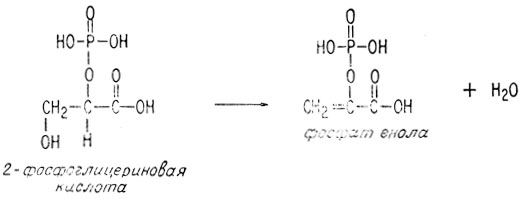
Образующийся в результате такой дегидратации ненасыщенный фосфат богат энергией, поскольку его электронная структура аналогична электронной структуре ангидрида. Он обладает также способностью превращать АДФ в АТФ.
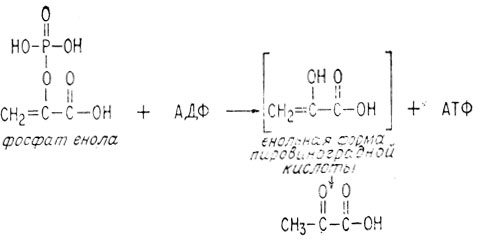
Таким образом, в итоге метаболизма одна молекула глюкозы, Две молекулы фосфорной кислоты и две молекулы АДФ превращаются в две молекулы пировиноградной кислоты и две молекулы АТФ. При этом также восстанавливаются две молекулы НАД+.
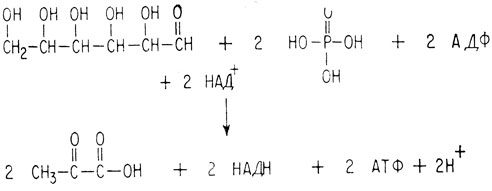
Суммарная реакция анаэробного метаболизма
Молекула глюкозы содержит пять одинарных связей и одну двойную связь углерод - кислород. Две молекулы пировиноградной кислоты содержат в сумме десять связей углерод - кислород, по пять в каждой молекуле. Таким образом, превращение глюкозы в пировиноградную кислоту является окислением, поскольку в образующихся молекулах связей углерод - кислород больше, чем в исходной молекуле. В результате этого окисления образуется энергия, которую организм накапливает и хранит в виде богатых энергией молекул АТФ. Следовательно, этот "склад энергии" служит для снабжения организма по мере надобности энергией, необходимой для поддержания жизни, роста и воспроизводства.
Для сохранения жизни необходим какой-то источник энергии, и первичные организмы, появившиеся на Земле, получали эту энергию в результате превращения органических молекул, таких, как глюкоза, синтезируемых абиогенным путем в окружающей среде. Таким образом, сложилось положение, при котором эти организмы постепенно сокращали свои источники питания, в то время как единственным источником пополнения, который в конце концов тоже имел предел, являлся остававшийся в атмосфере метан. Чтобы жизнь на Земле продолжалась, должен был быть обеспечен какой-то другой источник питательных молекул. Идеальным было бы, если бы этот новый источник был неиссякаем. Поскольку потребление питательных молекул происходило за счет окисления, возмещаться они должны были восстановительным путем. В противном случае в конце концов все органические молекулы в мире оказались бы окисленными в результате стадии питания. Атмосферный водород в качестве восстанавливающего агента долговременного действия был малопригоден, так как из-за малой массы он непрерывно улетучивался в космическое пространство, а образование его в результате фотолиза метана в конце концов прекратилось бы с исчерпанием метана. Создание дополнительного механизма восстановительного метаболизма, включающего, к примеру, превращение пировиноградной кислоты снова в глюкозу, невозможно, поскольку такого типа восстановление требует затраты энергии, а всякий процесс метаболизма должен поставлять энергию для поддержания жизни.
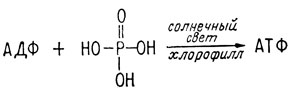
Таким образом, проблема заключалась в том, чтобы найти неисчерпаемый источник энергии и как-то использовать этот источник для восстановления сильно окисленных углеродных соединений до такой степени, чтобы они могли служить питательными веществами. Наиболее доступным природным источником энергии для этих целей являлось солнце, энергия которого уже использовалась для расщепления молекул первичной атмосферы на реакционноспособные фрагменты, которые, соединяясь, образовали формальдегид и цианистый водород. Нужно было найти способ уловить часть энергии, излучаемой солнцем, превратить ее в химическую форму и использовать эту энергию, например, для превращения АДФ в АТФ. Затем АТФ могла бы служить для поддержания необходимых восстановительных реакций. И этот способ был найден.
Живые организмы выработали процесс фотосинтеза, в ходе которого энергия солнца использовалась для образования АТФ, а АТФ в свою очередь расходовалась для проведения реакций восстановления. Первая стадия процесса фотосинтеза - поглощение солнечного света - осуществляется при помощи молекулы, называемой хлорофиллом, являющейся производным порфирина.
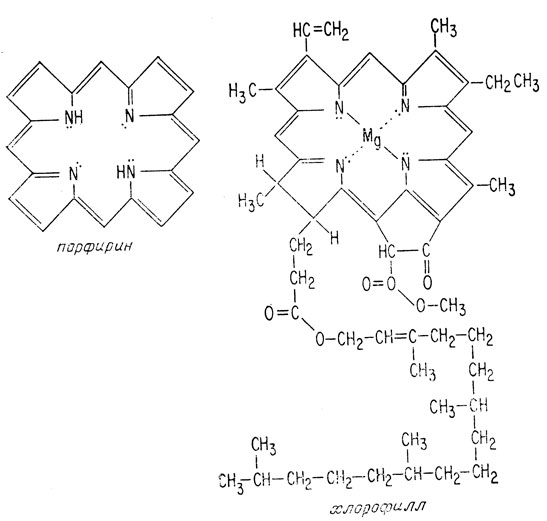
Производные порфирина, имевшие добиологическое происхождение, вероятно, присутствовали в первичном океане. Они были найдены в нефти, которая могла образоваться неорганическим путем. В результате недавно проведенных опытов, моделирующих добиологический синтез, получены также молекулы, напоминающие порфирин. Таким образом, в распоряжении первых живых организмов, вероятно, были молекулы такого типа.
Порфирин и производные порфирина содержат длинную последовательность чередующихся простых и двойных связей, называемую сопряженными двойными связями. Когда на такую молекулу попадает квант света, он возбуждает ее, переводя электроны на орбитали, обладающие более высокой энергией. Такая молекула называется фотовозбужденной.
В результате возвращения возбужденных электронов на свои исходные орбитали молекула высвобождает энергию и при соответствующих условиях эта энергия может быть использована для осуществления химической реакции. Именно это происходит с фотовозбужденным хлорофиллом. Он поставляет энергию, необходимую для превращения АДФ в АТФ.
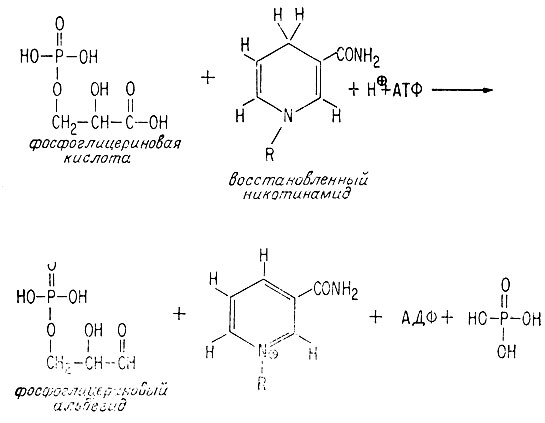
Организм может использовать энергию образовавшейся молекулы АТФ для восстановления окисленной молекулы, например фосфоглицериновой кислоты до молекулы, находящейся на уровне окисления углевода, например фосфоглицеральдегида. Обе эти молекулы - промежуточные соединения, образующиеся по ходу метаболизма при анаэробном окислении глюкозы в пировиноградную кислоту. Восстановителем служит восстановленный никотинамид, который также, как мы видели ранее, участвует в схеме анаэробного окисления глюкозы.
Значение процесса фотосинтеза состоит в том, что живой организм выработал способность потреблять энергию солнца, создавая для себя молекулы пищи. На пути эволюции это был колоссальный шаг вперед по сравнению с организмом, не использующим процесс фотосинтеза и зависящим от окружающей среды, которая поставляет ему предварительно образованные молекулы пищи.
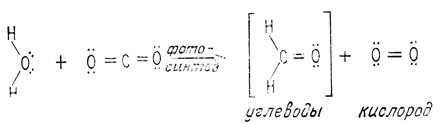
Способность восстанавливать углеродные соединения с помощью солнечной энергии позволила появившемуся живому организму усваивать двуокись углерода, возможно в виде иона карбоната или бикарбоната, из окружающего первичного океана и использовать эту двуокись углерода в качестве источника атомов углерода для образования молекул Сахаров и других питательных молекул. Эта реакция составляет сущность процесса фотосинтеза. В зеленых растениях при фотосинтезе происходит восстановление двуокиси углерода до альдегида (углевода), при котором вода служит первичным источником атомов водорода, а в атмосферу выделяется высвобождаемый при таком восстановлении кислород. Так же как и при анаэробном метаболизме (см. разд. "Первичный метаболизм"), каждая реакция, входящая в процесс фотосинтеза, требует участия строго специфического фермента.
Хотя этот процесс выглядит простым, на деле он исключительно сложен. Лишь недавно, после многолетних исследований, наконец в основном установлены этапы превращения атома углерода. Некоторые детали процесса все еще остаются невыясненными, например каким образом фотовозбужденный хлорофилл образует АТФ из АДФ и фосфата. Молекула двуокиси углерода или ион бикарбоната, попадая в клетку, где происходит фотосинтез, реагирует с пятиатомным сахаром (дифосфатом рибулозы), образуя две молекулы фосфоглицериновой кислоты. На это требуется всего несколько секунд.
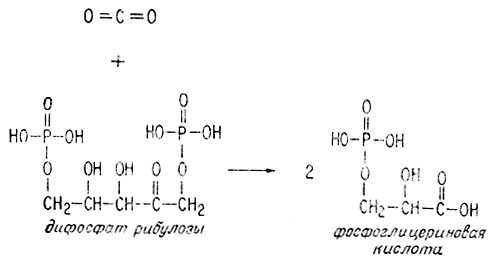
Как это происходит - не вполне ясно, по-видимому, процесс состоит из нескольких следующих стадий:
1) изомеризации дифосфата рибулозы
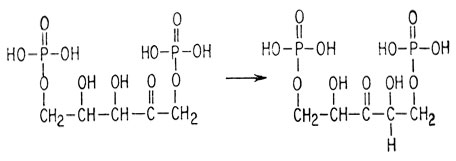
2) образования аниона сахара
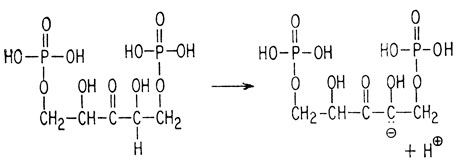
[Этот карбанион стабилизуется за счет енолизации (см. приложение).]
3) нуклеофильной атаки двуокиси углерода анионом
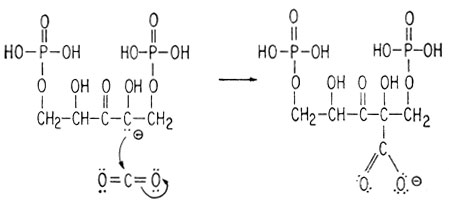
и 4) гидролитического расщепления (нуклеофильная атака молекулой воды)
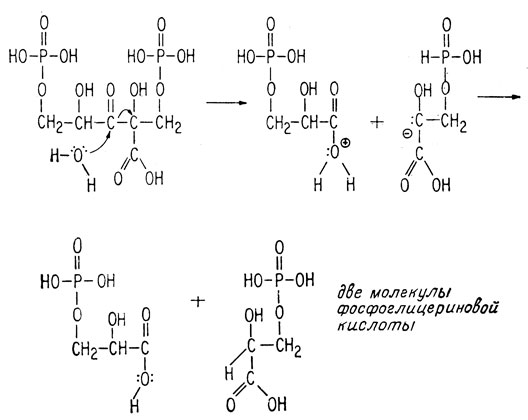
Эта гипотетическая схема подтверждается наблюдениями, сделанными в лаборатории Мелвина Калвина*, согласно которым атом углерода радиоактивной двуокиси углерода 14CO2 сперва появляется в карбоксильной группе фосфоглицериновой кислоты.
* (За пионерскую работу по установлению пути превращения углерода в процессе фотосинтеза профессору Калвину была присуждена Нобелевская премия по химии за 1961 г.)

Как было показано ранее, фосфоглицериновая кислота восстанавливается до фосфоглицеральдегида под действием АТФ и восстановленного никотинамида. Образовавшийся глицеральдегид может продолжать поддерживать процесс анаэробного метаболизма, идущего через образование фруктозы и глюкозы и заканчивающегося образованием запаса пищи в виде крахмала или в виде целлюлозы, откладывающейся в стенке клетки; как крахмал, так и целлюлоза являются полисахаридами.
Фотосинтез фактически является циклическим процессом. Приводимый в действие солнечным светом, каждый цикл включает поглощение трех молекул двуокиси углерода и образование одной молекулы фосфоглицериновой кислоты (рис. 2.2).
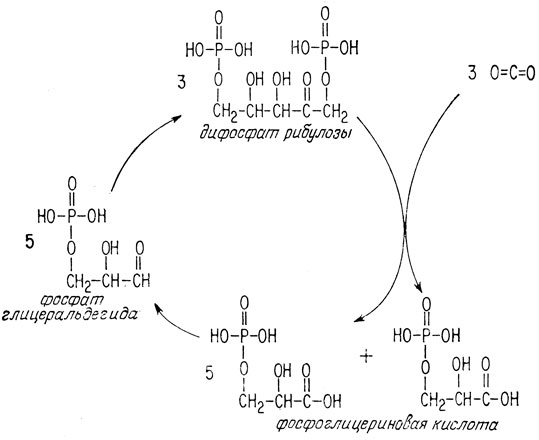
Рис. 2.2. Цикл фотосинтеза. В каждом цикле поглощаются три молекулы двуокиси углерода и образуется одна молекула фосфоглицериновой кислоты
Цикл начинают три молекулы дифосфата рибулозы. Они присоединяют три молекулы двуокиси углерода, образуя шесть молекул фосфоглицериновой кислоты. Пять из этих молекул остаются в цикле и восстанавливаются до фосфата глицеральдегида. Затем образовавшиеся пять молекул фосфата глицеральдегида взаимодействуют друг с другом, вновь образуя исходные три молекулы дифосфата рибулозы. Этим завершается один оборот цикла, и три молекулы дифосфата рибулозы начинают другой оборот, поглощая еще три молекулы двуокиси углерода, и т. д.
Первые две трети цикла (теперь мы будем рассматривать его, не упоминая фосфатные группы) - от рибулозы до глицеральдегида - уже были обсуждены. Последняя треть - превращение глицеральдегида опять в рибулозу - даже при записи в скелетной форме показывает, что атом углерода проходит чрезвычайно сложный путь превращения. Участвующие в этом процессе углеводы обозначены как триозы, тетрозы, пентозы и т. д. в соответствии с количеством содержащихся в них атомов углерода (рис. 2.3).
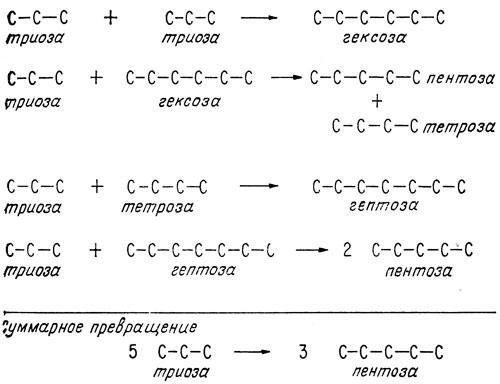
Рис. 2.3. Суммарная запись реакций, участвующих во взаимопревращениях углеводов и составляющих часть цикла фотосинтеза. Пять молекул глицеральдегида (триоза) превращаются в три молекулы рибулозы (пентоза)
Следует еще раз подчеркнуть, что все реакции, входящие в цикл фотосинтеза, катализируют ферменты. В целом цикл требует затраты энергии, поскольку он имеет восстановительный характер; эта энергия поставляется АТФ, а она образуется фотовозбужденным хлорофиллом. Фотовозбужденный хлорофилл поставляет также энергию, необходимую для восстановления НАД+ до НАДН, который используется в цикле в качестве восстанавливающего агента. В зеленых растениях атомы водорода, переходящие в процессе этого восстановления к НАД+, поставляет вода.
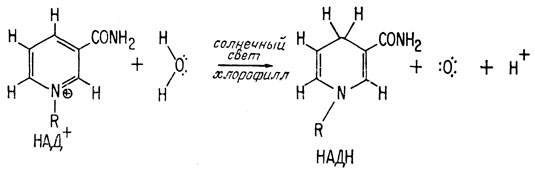
Наряду с получением восстановленного никотинамида, необходимого для образования молекул пищи, при этой реакции освобождается кислород, и, таким образом, она явилась источником образования богатой кислородом атмосферы, существующей в настоящее время. Итак, именно процесс фотосинтеза в первичном океане привел не только к созданию нового источника питания, но и сделал доступным кислород для существовавших в то время живых форм. Присутствие кислорода в свою очередь привело к следующей стадии совершенствования жизни на Земле - созданию аэробного метаболизма.
Этот новый механизм аэробного обмена веществ не заменил собой старый анаэробный метаболизм. Скорее, следует сказать, что новый механизм наложился на старый, дополнив и улучшив его. Аэробный метаболизм вступает в дело там, где перестает действовать анаэробный метаболизм. Этот процесс начинается с конечного продукта анаэробного метаболизма - пировиноградной кислоты - и продолжает окислительное разложение до самого конца - до образования двуокиси углерода и воды (рис. 2.4). Значение этого процесса состоит в том, что им завершается третье, последнее звено жизненного цикла, что теоретически делает возможным бесконечное продолжение жизни на Земле, по крайней мере до тех пор, пока светит Солнце.
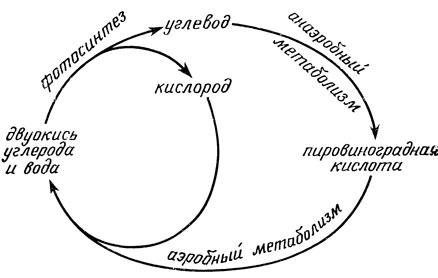
Рис. 2.4. Жизненный цикл
Создание механизма аэробного метаболизма принесло также существенные практические выгоды. При новом процессе метаболизма стало получаться гораздо большее число молекул АТФ, чем при анаэробном метаболизме. При анаэробном метаболизме молекула глюкозы окисляется до двух молекул пировиноградной кислоты и образуются две молекулы АТФ. При аэробном метаболизме происходит дальнейшее окисление двух молекул пировиноградной кислоты до двуокиси углерода и воды и образуется 30 молекул АТФ. Аэробная схема также включает окисление 2 молекул НАДН, полученных анаэробным путем. При этом получается еще 6 молекул АТФ, что в итоге приводит к образованию 38 молекул АТФ на каждую молекулу глюкозы, полностью окислившуюся до двуокиси углерода и воды (рис. 2.5).

Рис. 2.5. Суммарная реакция клеточного дыхания
Аэробный метаболизм представляет собой процесс, состоящий из двух частей. В первой его части пировиноградная кислота окисляется до двуокиси углерода, а НАД+ восстанавливается до НАДН и Н+. Эта часть называется циклом образования лимонной кислоты, поскольку процесс является циклическим, а лимонная кислота образуется в качестве промежуточного соединения. Во второй части НАДН+ окисляется опять до НАД+, кислород восстанавливается до воды и образуется АТФ. Эта часть процесса называется дыхательной цепью.
Перед вступлением в цикл образования лимонной кислоты пировиноградная кислота претерпевает окислительное декарбоксилирование. Эта реакция, как и большинство биологических реакций, катализируется ферментом (гл. 3). В действительности

в ходе этой реакции свободная уксусная кислота не образуется. Образуется замещенное у атома серы производное (тиоэфир) уксусной кислоты, называемое ацетильным коферментом А (гл. 10). Этот тиоэфир включается в цикл образования лимонной кислоты, присоединяясь к щавелевоуксусной кислоте, в результате чего получается лимонная кислота.
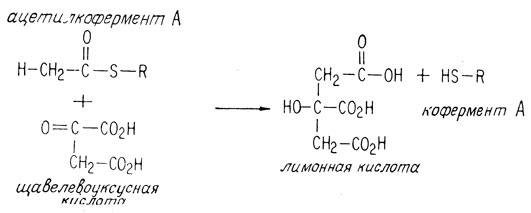
Затем лимонная кислота изомеризуется под влиянием фермента в изолимонную кислоту.

После этого изолимонная кислота одновременно окисляется и декарбоксилируется, образуя α-кетоглутаровую кислоту и вторую молекулу двуокиси углерода.
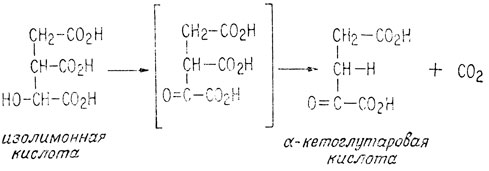
Для того чтобы в процесс окисления оказались вовлеченными все три углеродных атома пировиноградной кислоты, должна образоваться только одна молекула двуокиси углерода. Это осуществляется при окислительном декарбоксилировании образовавшейся молекулы α-кетоглутаровой кислоты, аналогичном окислительному декарбоксилированию пировиноградной кислоты.
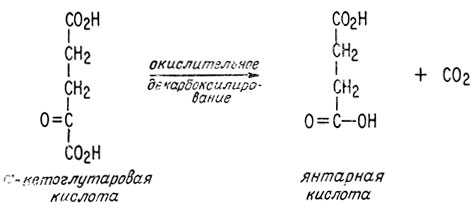
В завершающей стадии цикла просто регенерируется щавелевоуксусная кислота за счет окисления янтарной кислоты.
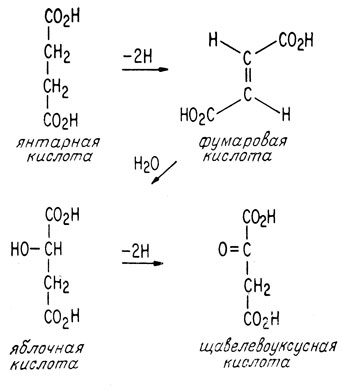
Непосредственно в цикле образования лимонной кислоты получается только одна молекула АТФ. Это происходит при окислительном декарбоксилировании α-кетоглутаровой кислоты. Все остальные молекулы АТФ, получаемые в процессе аэробного метаболизма, образуются в дыхательной цепи. Основная роль цикла образования лимонной кислоты заключается в снабжении дыхательной цепи атомами водорода в виде НАДН и Н+. При каждом повторении цикла образования лимонной кислоты три молекулы НАД+ восстанавливаются до НАДН. Первая молекула восстанавливается при окислительном декарбоксилировании пировиноградной кислоты, вторая - при окислительном декарбоксилировании α-кетоглутаровой кислоты, а третья молекула восстанавливается при дегидрировании яблочной кислоты (рис. 2.6).
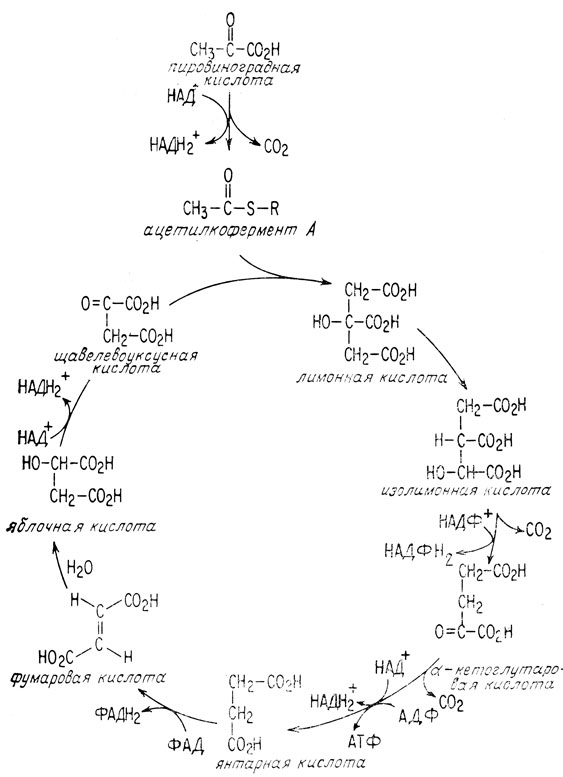
Рис. 2.6. Цикл образования лимонной кислоты
Кроме того, при окислении и декарбоксилировании изолимонной кислоты восстанавливается молекула никотинамидадениндинуклеотидфосфата (НАДФ+). Эта молекула отличается от НАД+ тем, что она содержит три, а не два остатка фосфорной кислоты. Никотинамидные части обеих молекул выполняют окислительно-восстановительные функции аналогичным образом.
Когда янтарная кислота теряет два атома водорода, превращаясь в фумаровую кислоту, окисляющим агентом является не НАД+, а ФАД (флавинадениндинуклеотид); молекула ФАД подобна молекуле НАД+, за исключением того, что вместо окислительно-восстановительной системы никотинамида она содержит окислительно-восстановительную систему флавина. В обеих этих системах окислительно-восстановительные реакции происходят на атомах азота, а боковая группа R одинакова для обеих молекул.
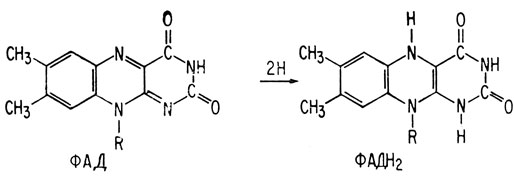
Восстановление флавинадениндинуклеотида
Rфлавина - это аденин-рибоза-фосфат-фосфат-рибоза-флавин; Rникотинамида - это аденин-рибоза-фосфат-фосфат-рибоза-никотинамид.
Таким образом, каждое повторение цикла образования лимонной кислоты приводит к восстановлению четырех молекул никотинамида и одной молекулы флавина. Каждая из этих восстановленных молекул поставляет в дыхательную цепь пару атомов водорода (рис. 2.7).

Рис. 2.7. Дыхательная цепь
Дыхательная цепь состоит из ряда одновременно протекающих окислительно-восстановительных реакций. Начинается эта цепь с окисления НАДН, а кончается восстановлением кислорода молекулами, называемыми цитохромами и участвующими в промежуточных стадиях. При каждом прохождении цепи образуются три молекулы АТФ.
Цитохромы, участвующие в дыхательной цепи, представляют собой гем-содержащие белки. Гем - это производное порфирина, содержащее ион железа. Когда цитохром находится в окисленном состоянии, он содержит ион трехвалентного железа (Fe3+). В восстановленном состоянии он содержит ион двухвалентного железа (Fe2+). Эти два состояния отличаются только на один электрон. Поскольку каждый атом водорода содержит один электрон, для
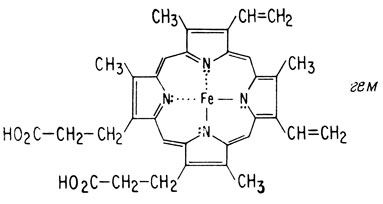
окисления одной молекулы ФАДН2 до ФАД требуются два гема, содержащих ион трехвалентного железа.
Молекула ФАДН2, получающаяся в цикле образования лимонной кислоты, вступает в дыхательную цепь, восстанавливая цитохром b3+ (рис. 2.7). Эта последовательность реакций идет без участия НАД+, и поэтому в результате появления в дыхательной цепи двух атомов водорода из ФАДН2 получаются только две молекулы АТФ. Этот дефицит компенсируется, однако, "лишней" молекулой АТФ, образуемой при окислительном декарбоксилировании α-кетоглутаровой кислоты. Таким образом, в результате каждого повторения цикла образования лимонной кислоты образуются четыре молекулы восстановленного никотинамида, одна молекула восстановленного флавина и одна молекула АТФ. Четыре пары атомов водорода от восстановленных молекул никотинамида образуют в дыхательной цепи двенадцать молекул АТФ, а два атома водорода от восстановленного флавина образуют еще две молекулы АТФ. Это (плюс еще одна молекула АТФ, получаемая непосредственно в цикле образования лимонной кислоты) составляет в сумме пятнадцать молекул АТФ на каждую молекулу пировиноградной кислоты, окисленную до двуокиси углерода и воды.
Анаэробное окисление молекулы глюкозы до двух молекул пировиноградной кислоты приводит к образованию двух молекул восстановленного никотинамида и двух молекул АТФ. С развитием аэробного метаболизма две молекулы восстановленного никотинамида смогли передавать выделяемые ими водородные атомы в дыхательную цепь для образования еще шести молекул АТФ. Сочетание анаэробного и аэробного метаболизмов приводит, таким образом, к образованию в сумме 38 молекул АТФ на каждую молекулу глюкозы, окислившуюся до двуокиси углерода и воды.
Однако истинным чудом этого изобретения природы является высокая степень эффективности, достигнутая при улавливании и хранении энергии окисления. Для превращения моля АДФ в АТФ требуется около 12 ккал. При сгорании глюкозы до двуокиси углерода и воды высвобождается около 690 ккал/моль. В 38 молях АТФ улавливается и хранится 456 ккал; следовательно, к. п. д. этого превращения составляет 66%. Обычно паровые силовые установки имеют примерно вдвое меньший к. п. д.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'