2.5. Транспорт веществ при участии переносчиков
Глицерин проникает в эритроциты человека или крысы значительно быстрее, чем в эритроциты других млекопитающих, в том числе в эритроциты свиньи, овцы и коровы. Имеются данные о том, что перенос глицерина через мембрану эритроцитов человека и крысы протекает по специальному механизму. Кинетика этого процесса сильно отличается от обычного пассивного транспорта. Перенос глицерина через мембрану эритроцитов человека и крысы характеризуется очень низкой энергией активации, а скорость этого процесса не имеет ярко выраженной температурной зависимости. Представляется весьма вероятным, что механизм переноса глицерина через мембрану не связан с полной дегидратацией. Это означает, что путь глицерина не проходит непосредственно через углеводородный слой липида, образующего мембрану. Такой механизм транспорта веществ через мембрану предусматривает участие специальных переносчиков белковой природы.
Аналогичным закономерностям подчиняется транспорт глюкозы в эритроциты. Глюкоза имеет пять гидроксильных групп, способных образовывать водородные связи. Теоретически энергия активации процесса переноса глюкозы из водной фазы в гидрофобную область мембраны должна составлять около 80 кДж/моль, однако величина энергии активации, полученная в экспериментах, равна приблизительно 16 кДж/моль. Вследствие этого перенос глюкозы через мембрану эритроцитов происходит примерно в 10 000 раз быстрее, чем это можно было бы ожидать при пассивном транспорте.
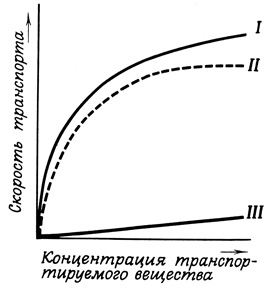
Рис. 29. Различие между пассивным (III) транспортом и транспортом при участии переносчиков (II). График I характеризует общую скорость переноса
Транспорт веществ при участии переносчиков характеризуется некоторыми чертами, позволяющими отличать его от пассивного транспорта (рис. 29). Прежде всего он очень специфичен. Система, осуществляющая такой специфический (при участии переносчиков) транспорт веществ через мембрану, как правило, имеет ограниченную мощность, проявляя тенденцию к насыщению. При возрастании химического или электрохимического градиента в такой системе скорость переноса должна приближаться к максимальной величине и оставаться на этом уровне* при условии, конечно, что концентрация транспортируемого вещества может быть увеличена до соответствующих пределов.
* (Следует заметить, что в реальной системе общий поток через мембрану всегда включает компонент, отражающий скорость пассивного транспорта. Это обстоятельство надо постоянно иметь в виду при рассмотрении феномена насыщения.)
Коэффициент простой диффузии и уравнение Нернста - Планка для пассивной проницаемости не могут адекватно описать кинетику транспорта веществ через мембрану при участии переносчиков. Успешные попытки анализа кинетических закономерностей такой системы основаны на предположении, что специфическое взаимодействие транспортируемых молекул с некими компонентами мембраны, характеризующееся константой сродства* Km, является наиболее важным фактором всего процесса. Однонаправленный поток вещества через мембрану описывается выражением, аналогичным уравнению Михаэлиса - Ментен:
* (Константа сродства может быть выражена как концентрация проникающего вещества, при которой J=Jмакс/2.)
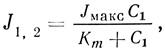
где J1,2 представляет собой поток вещества из отсека 1 в отсек 2, Jмакс - максимальная скорость потока, т. е. значение скорости в условиях полного насыщения переносчика, и C1 - концентрация транспортируемого вещества в отсеке 1. Для результирующего потока вещества через мембрану выражение будет иметь вид
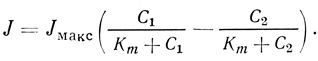
В большинстве систем, характеризующихся транспортом веществ при участии переносчиков, трансмембранный электрохимический градиент транспортируемого вещества поддерживается благодаря модификации этого вещества или удаления его из второго отсека. Это означает, что скорость переноса с хорошим приближением можно определить по формуле

Формальное сходство этого выражения с уравнением ферментативной кинетики позволяет использовать некоторые математические приемы обработки экспериментальных результатов, применяемые в ферментативной кинетике, для определения Jмакс и Кm, а также для характеристики ингибиторов транспорта (рис. 30).
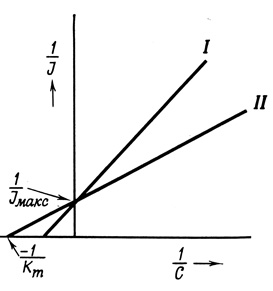
Рис. 30. Использование метода Лайнуивера - Бэрка для изучения конкурентного ингибирования процесса переноса через мембраны. I - добавление конкурентного ингибитора; II - без ингибитора
Ярким примером явления насыщения служит повторная адсорбция различных веществ из клубочкового фильтрата клетками, выстилающими почечные канальцы (рис. 31). Например, глюкоза будет полностью реадсорбирована клетками канальцев только в том случае, если ее общее содержание в клубочковом фильтрате не превышает уровня насыщения системы переносчиков в клеточной мембране. Эта точка насыщения при нормальном кровотоке в почках соответствует концентрации глюкозы в крови приблизительно 180 мг/мл (почечный порог для глюкозы). Если концентрация глюкозы в крови превышает эту величину, то избыток глюкозы остается в фильтрате и выделяется с мочой (глюкозурия). Это можно наблюдать при сахарном диабете, характерным признаком которого является высокий уровень содержания глюкозы в крови. Причина столь высокого содержания глюкозы в крови больных сахарным диабетом кроется в неспособности метаболической системы этих больных эффективно удалять глюкозу из циркулирующей крови. Вместе с тем если в моче обнаруживается глюкоза, а концентрация ее в крови ниже нормального почечного порога, то, следовательно, нарушен транспорт глюкозы в почках. В моче обнаруживаются и другие вещества, реадсорбируемые в почках при участии специальной системы переносчиков, если концентрация этих веществ в крови превышает почечный порог.
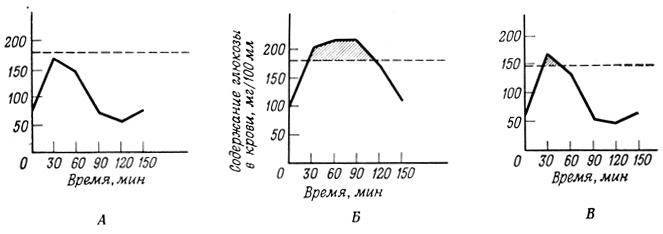
Рис. 31. Проба на толерантность к глюкозе. В момент времени 0 испытуемым давали 50 г глюкозы per os. А - контроль; Б - больные диабетом. Пунктирной линией в этих двух случаях обозначен нормальной почечный порог; В - больные с почечной глюкозурией. Пунктирной линией в этом случае обозначен ненормальный почечный порог. Заштрихованные участки соответствуют выделению глюкозы с мочой (глюкозурии)
Для некоторых систем (например, систем транспорта ряда веществ в эритроциты человека) практически нереально достигнуть требуемой для насыщения концентрации транспортируемого вещества и в то же время не нарушить целостности мембраны. В таком случае для выяснения механизма транспорта веществ через мембрану невозможно использовать критерий "насыщения".
Переносчики обладают, как правило, высокой избирательностью при взаимодействии с различными типами транспортируемых молекул (они, например, различают L- и D-изомеры некоторых сахаров и аминокислот), однако часто они транспортируют близкие по структуре соединения. Эти соединения, следовательно, конкурируют друг с другом за место связывания на переносчике. Так, минимальное требование, которому должна отвечать структура молекулы для того, чтобы система транспорта сахара в кишечнике млекопитающих могла узнать ее, - это наличие шестиуглеродного пиранозного скелета с гидроксильной группой у второго атома углерода.
Таким образом, D-глюкоза и D-галактоза (рис. 32), различающиеся только ориентацией одной гидроксильной группы и удовлетворяющие вышеуказанным требованиям, конкурируют друг с другом за системы транспорта сахаров в некоторых клетках. В этом случае оба конкурирующих соединения проникают через мембрану, тогда как большое число конкурентных ингибиторов просто блокирует транспортные системы, но не проходит через мембрану. Такие ингибиторы затрудняют доступ транспортируемых молекул к переносчику. Флоридзин (рис. 33), например, конкурентно ингибирует транспорт глюкозы в некоторые клетки, не проникая при этом через клеточную мембрану. Если несколько соединений переносится через мембрану одной и той же транспортной системой, это не обязательно означает, что они в равной степени конкурируют за связывание с переносчиком. Более того, системы переносчиков в различных клетках или тканях могут отличаться на несколько порядков по своему сродству к одной и той же группе соединений.
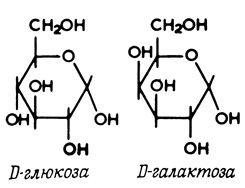
Рис. 32. D-глюкоза и D-галактоза, конкурирующие между собой за связывание с переносчиком. Следует отметить, однако, что каждое из этих соединений не ингибирует транспорт другого
Помимо специфических ингибиторов, имеющих структурное сходство с транспортируемыми молекулами, встречаются также соединения, сильно отличающиеся по своей структуре от нормальных метаболитов и в то же время эффективно воздействующие на транспортные системы. Уабаин (рис. 33) в исключительно низких концентрациях подавляет сопряженный транспорт натрия и калия, а инсулин стимулирует транспорт глюкозы в некоторые типы клеток млекопитающих.
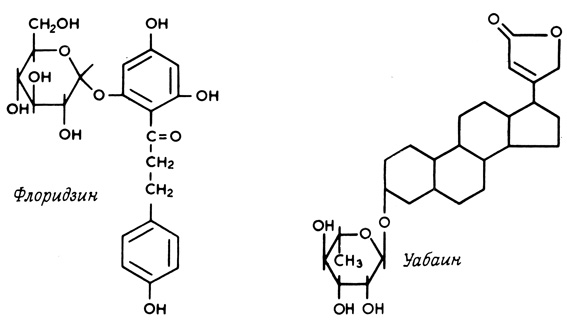
Рис. 33. Структурные формулы ингибиторов опосредованного (при участии переносчиков) транспорта - флоридзина и уабаина (строфантина G)
Еще один подход, позволяющий идентифицировать механизм транспорта веществ через мембрану при участии переносчиков, заключается в изучении зависимости скорости этого процесса от pH (рис. 34), а также неконкурентного ингибирования транспорта ионами тяжелых металлов, тиоловыми соединениями, динитрофторбензолом и т. д., т. е. в исследовании влияния тех факторов, которые изменяют активность белковых молекул. Например, транспорт глицерина при участии переносчиков в эритроциты человека или крысы, протекающий с высокой эффективностью при pH 7,0, резко замедляется при понижении pH системы ниже 6,0 или при обработке клеток низкими концентрациями солей меди (<0,1 мМ CuSO4). Чувствительность транспортных систем к низким концентрациям ингибиторов рассматривается как доказательство того, что каждая система занимает лишь небольшую часть поверхности всей мембраны.
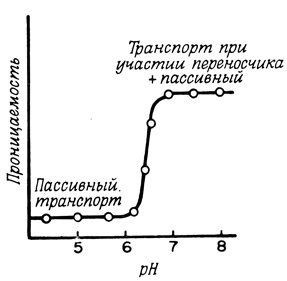
Рис. 34. Зависимость пассивного транспорта и транспорта при участии переносчиков от величины pH
Обменная диффузия и противотранспорт - явления, характерные для системы транспорта с участием переносчиков. Оба процесса можно наблюдать экспериментально. Под термином "обменная диффузия" подразумевают быстрый трансмембранный обмен молекул одного и того же вещества при участии переносчиков. Когда концентрация транспортируемого вещества высока по обе стороны мембраны и система находится в равновесии, суммарный поток вещества через мембрану равен нулю. Если теперь в такую систему по одну сторону мембраны добавить небольшое количество того же самого вещества, несущего радиоактивную метку, то будет наблюдаться быстрый поток меченых молекул через мембрану в обмен на немеченые молекулы (рис. 35). Описанный эксперимент показывает, что даже в условиях низкого суммарного потока вещества через мембрану существует значительный трансмембранный обмен молекул этого вещества. Обменная диффузия подчиняется кинетике Михаэлиса - Ментен и характеризуется явлением насыщения, поэтому ее можно рассматривать как транспорт с участием переносчиков.
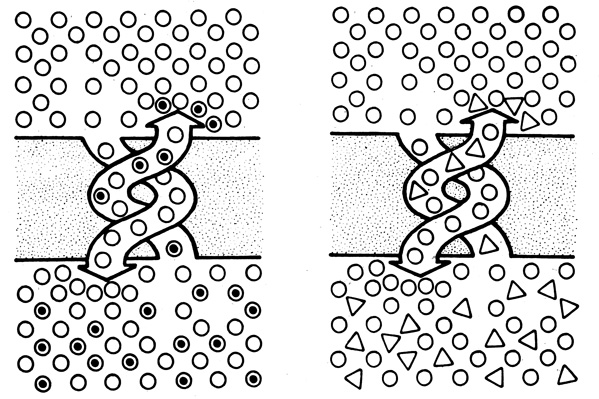
Рис. 35. Обменная диффузия (слева) и антипорт (справа) Кружками с черной точкой внутри обозначено меченое вещество; белые кружки и треугольники - различные транспортируемые вещества
Противотранспорт происходит в том случае, если в систему, содержащую один переносчик, способный транспортировать два вещества А и Б, и уравновешенную по веществу A (JA1,2=JA2,1), добавляют вещество Б по одну сторону мембраны (например, в отсек 1). Конкуренция вещества А с веществом Б за переносчик приведет к тому, что поток JA1,2 уменьшится, тогда как поток JA1,2 останется без изменений. В результате вещество А будет накапливаться в отсеке 1, несмотря на то что исходная его концентрация по обе стороны мембраны была одинакова.
В сравнительно небольшом числе систем транспорт одного вещества через мембрану зависит от присутствия другого вещества (рис. 36). Это явление совместного переноса веществ через мембрану следует также отнести к транспорту при участии переносчиков. Примером такой системы транспорта служит совместный (одновременный) перенос в противоположных направлениях (часто называемый антипортом) Na+ и К+ через плазматические мембраны, большого числа клеток, а также АТФ и АДФ через внутреннюю митохондриальную мембрану или совместный перенос в одном и том же направлении (симпорт) Na+ и некоторых сахаров, а также Na+ и некоторых аминокислот через мембрану эпителиальных клеток почки и кишечника.
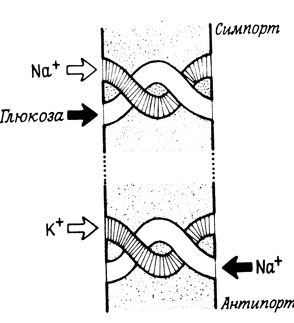
Рис. 36. Различные системы совместного транспорта
Избирайте себе милую индивидуалку и пишите ей в любое время дня, чтобы доболтаться о свидании. Всегда очаровательные проститутки уже готовы прикатить к вам в гости и усладиться обольстительным трахом без запретов.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'