Часть V. Применение телеметрии при изучении физиологии животных и человека
Телеметрическая регистрация физиологических параметров у находящихся в полете птиц (Э. Эльяссен)
Мы очень мало знаем о физиологии птиц в обычном полете. Так как полет связан с большим физическим напряжением, было бы интересно знать, какие требования предъявляет он к различным физиологическим функциям - энергетическому обмену веществ, терморегуляции и кровообращению.
При изучении кровообращения, точнее сердечно-сосудистых реакций при полете, возникают два основных вопроса: какова частота сердечных сокращений и кровяное давление сразу же после взлета птицы и через некоторое время после взлета.
Для измерения частоты сердечных сокращений и кровяного давления нужно использовать телеметрическую систему, состоящую из передатчика и приемника. Вес и размеры передатчика должны быть такими, чтобы не нарушить движения птицы в полете, и прикреплять его к птице следует таким образом, чтобы не сместить ее центр тяжести. Возможны два подхода: внутреннее или наружное крепление передатчика. Можно сделать передатчик минимальным по размерам и весу и ввести его в сердце или в сосуд. Однако это кажется мне довольно трудной операцией по следующим причинам: сердце птицы и ее самые крупные артериальные сосуды расположены так, что ввести в них передатчик крайне трудно. По очевидным причинам невозможно пройти через грудину, и остается единственный путь - обойти ее спереди. Но тогда можно повредить передние воздушные мешки.
Я хочу подчеркнуть, что необходимо принимать во внимание биологические особенности объекта, к которому мы прикрепляем передатчик. Это очень важный фактор. При обсуждении различных технических проблем мы никогда не должны забывать биологию.
Действительно, в известных пределах вопрос состоит не в том, чтобы сделать передатчик минимальным по размерам и весу, а в том, чтобы наилучшим образом приспособить его для определенных целей.
В настоящем докладе обсуждается способ внешнего крепления прибора для дистанционного измерения частоты сердечных сокращений и кровяного давления. Сначала дается общее описание аппаратуры, включая описание статической и динамической калибровки всей экспериментальной установки. Затем описывается способ измерения и приводятся некоторые предварительные результаты.
Технические особенности аппаратуры
Вес передатчика составляет около 40 г. Он включает в себя передающую систему, катетер и иглу для подкожных инъекций. Если использовать его для работы с большими птицами, например с домашними утками (вес около 1,5 кг) и морскими чайками (вес около 2 кг), то вес передатчика составляет всего 2-2,5% веса птицы.
Датчик представляет собой, по существу, конденсатор переменной емкости, включенный в настроенную схему генератора на транзисторе. Любое изменение давления в полости вызывает изменение емкости конденсатора и, следовательно, частоты генератора. Таким образом, мы получаем генератор с частотной модуляцией.
Приемная часть системы состоит из обычного приемника для частотно-модулированных сигналов и записывающего устройства.
Выходным сигналом приемника является напряжение постоянного тока, величина которого пропорциональна изменению частоты передатчика. Если предположить, что существует линейная зависимость между изменением частоты генератора и изменением давления, то выходная величина приемника, поступающая на записывающее устройство, также пропорциональна давлению в полости. В приемнике используется обычный трехточечный генератор с индуктивной обратной связью (схема Гартлея), в котором в качестве усилителя используется полупроводниковый тетрод. Частота генератора (около 90 мгц) определяется индуктивностью L и емкостью С. Для уменьшения длины антенны передатчика, которая не должна превышать 3 м частота должна быть по возможности большой. Антенна соединяется с передатчиком индуктивно, причем коэффициент связи делается минимальным, чтобы уменьшить влияние движений антенны на частоту генератора. Для этой цели можно также использовать для развязки антенны и генератора буферный каскад на транзисторе.
На фиг. 1 изображена схема датчика давления. Приведены также его основные размеры и указаны материалы, применявшиеся для различных частей датчика.
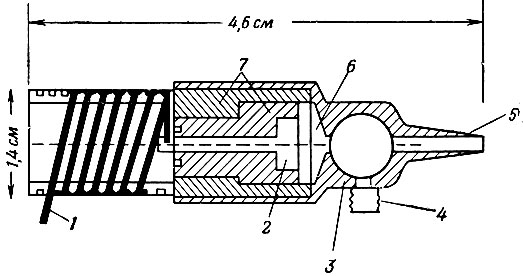
Фиг. 1. Эскиз датчика давления. 1 - катушка (индуктивность L); 2 - медная пластинка; 3 - алюминий; 4 - присоединение калибрующей системы; 5 - присоединение катетера; 6 - медная мембрана толщиной 0,1 мм; 7 - эбонит
Мембрана и медная пластинка образуют переменную емкость генератора. В датчике находится также индуктивность L, для того чтобы настраиваемая схема генератора была механически устойчивой.
Для определения чувствительности датчика (изменение емкости на единицу изменения давления) на него подавались известные по величине давления, а изменение емкости измерялось емкостным мостом. Полученная зависимость (фиг. 2) имеет линейный характер (для данного диапазона давлений). Чувствительность датчика составляет 1 пф на 1000 мм вод. ст.
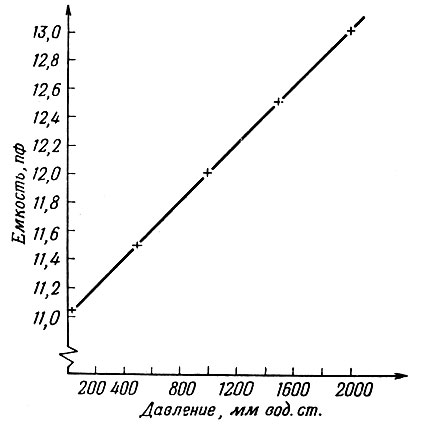
Фиг. 2. Характеристика датчика давления (см. текст)
При измерении давления в сосудах возможны явления брадикардии и тахикардии. В связи с этим необходимо знать амплитудно-частотную характеристику системы измерения давления, т. е. нужно определить динамическую характеристику этой системы.
Измерения проводились посредством стандартных электронных приборов и различных экспериментальных установок для диапазона низких и высоких частот. В первой установке использовались самописцы лаборатории Санборна, а во второй - электронно-лучевой осциллоскоп. Для обеих установок пришлось изготовить специальные приборы, с помощью которых получали синусоидальные изменения давления, необходимые при исследовании динамики.
При анализе датчика использовались частотные характеристики и переходные функции. Характеристики определялись путем сравнения реакции датчика с реакцией вспомогательной системы, состоящей из усилителя и манометра. Частота среза последней системы находилась далеко за пределами предполагаемого рабочего диапазона частот.
На второй экспериментальной установке для высоких частот исследовалась реакция датчика на колебания от 20 до 1500 рад/сек (3-250 гц).
Для получения синусоидальных изменений давления в наполненном водой датчике и контрольном манометре с успехом использовались звуковые колебания, возникавшие при перемещении постоянного магнита в электромагнитном поле, величина которого регулировалась при помощи усилителя. Выходной сигнал этого усилителя мог непрерывно изменяться от нуля до 190 в, что позволяло в широких пределах регулировать амплитуду давления. Такая регулировка давала возможность снимать характеристики либо при постоянной амплитуде выходного сигнала манометра вспомогательной системы, либо при постоянном выходном сигнале исследуемого датчика.
Результаты исследований, приведенные на фиг. 3, показали, что датчик имеет линейную характеристику в пределах ±2 дб до частоты 250 рад/сек (40 гц) и резонансный пик при 80 гц, т. е. далеко за максимально возможной частотой сердечных сокращений.
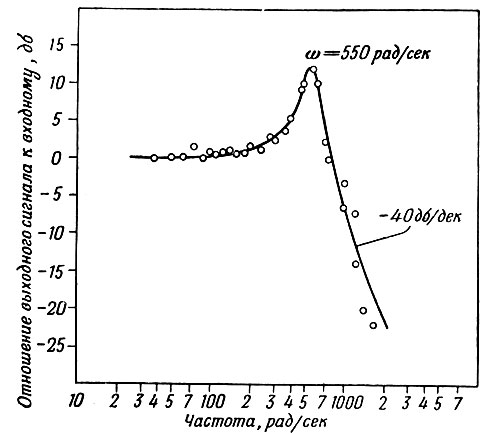
Фиг. 3. Частотная характеристика датчика давления (высокие частоты)
Характеристики на низкой частоте были необходимы для исследования датчика при работе с тонкой иглой для подкожных инъекций (внутренний диаметр 0,4 мм) и катетером такого же диаметра, имеющим длину 15 см.
Полученные результаты приведены на фиг. 4. Частота среза равна 8 гц. Сплошная линия на фиг. 4 близка к частотной характеристике фильтра нижних частот с единственной постоянной времени, равной 0,069 сек.
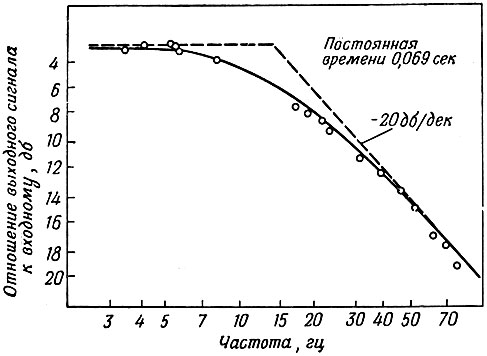
Фиг. 4. Частотная характеристика датчика давления (низкие частоты)
Полученная частота среза соответствует 500 сердечным сокращениям в минуту. Это вполне удовлетворительно для наших опытов с домашними утками и морскими чайками. Однако при опытах с меньшими по размерам птицами нужны другие приборы, в частности более короткий катетер с большим диаметром.
Результаты, полученные при анализе частотных характеристик, хорошо совпадают с результатами анализа переходных функций. Переходные функции определялись при подаче в систему скачка давления в 100 мм вод. ст. Реакции вспомогательной системы и датчика записывались самописцами в лаборатории Санборн. Для точного анализа ошибок переходного процесса применялась полулогарифмическая шкала.
На этом я закончу техническое описание системы. Добавлю лишь, что характеристики самого датчика существенно отличаются от характеристик всей измерительной системы. Это различие обусловлено, конечно, гидравлическим сопротивлением и емкостью соединительной системы, т. е. катетера и иглы для подкожных инъекций.
Способ телеизмерения
Передатчик прикрепляется по средней линии спины птицы (фиг. 5). Мы нашли целесообразным поместить всю систему в льняную ткань, оформленную в виде коробки с крышкой и пришитую к коже птицы.
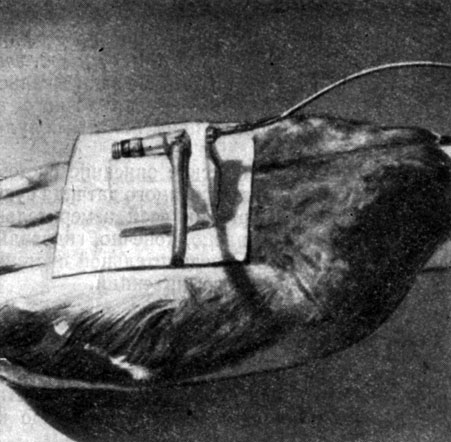
Фиг. 5. Передатчик, прикрепленный по средней линии спины птицы
До полета через резиновую трубку (видную на фиг. 5) и трехходовой кран в катетер подается гепарин. При этом кран повернут так, чтобы гепарин попал в катетер и сосудистую систему, но не смешивался с водой в датчике. Катетер через яремную вену вводился в правый желудочек.
Принимая во внимание довольно большой вес батарей передатчика, мы решили не крепить передатчик на уровне сердца, что обеспечило бы правильную нулевую линию при измерении давления. При наличии более легких батарей такое расположение аппаратуры получить нетрудно. В наших же опытах расстояние между уровнем сердца и уровнем датчика составляло (для утки) около 5 см.
Учитывая результаты опытов, можно сделать заключение, что нулевая линия как-то меняется во время полета. До того как этот недостаток будет устранен, можно считать мало существенными любые другие изменения в размещении аппаратуры на птице. Таким образом, при измерениях было получено только относительное давление, т. е. пульсовое, а не диастолическое и систолическое давление в сердце и артериях.
Опыты проводились в большом зале длиной 100 м и высотой 30 м. Благодаря этому мы могли ловить птиц после каждого опыта и проводить несколько опытов с одной и той же птицей.
Результаты опытов
На фиг. 6 приведены записи частоты сердечных сокращений и пульсового давления в правом желудочке перед полетом и во время полета птицы. Отрезок длиной 3 см по горизонтали соответствует 1 сек, а 2,6 мм по вертикали - калибровочному давлению 250 мм вод. ст.
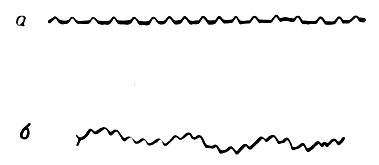
Фиг. 6. Частота сердечных сокращений и пульсовое давление (в правом желудочке) перед полетом (а) и во время полета (б) птицы
Частота сердечных сокращений составляла 300 ударов в минуту до полета и 360 ударов в минуту во время полета. Пульсовое давление было одинаковым до полета и во время него и составляло около 300 мм вод. ст.
Для сравнения на фиг. 7 приводится давление в правом желудочке той же птицы, полученное введением катетера и записанное при помощи датчика и усилителя, применяемых обычно при медицинских исследованиях. Здесь отрезок длиной 4 см по горизонтали соответствует 1 сек, а 5 мм по вертикали - калибровочному давлению 500 мм вод. ст. Полученные значения частоты сердечных сокращений и пульсового давления (соответственно 250 ударов в минуту и 350 мм вод. ст.) хорошо согласуются с данными телеметрической системы. Полученные данные показывают, что при полете птицы частота сердечных сокращений изменяется относительно мало, а пульсовое давление совсем не меняется (данные записаны после того, как птица пролетела 100 м).

Фиг. 7. Давление в правом желудочке утки (измерения проводились на той же утке, что и для фиг. 6)
На фиг. 8 показаны частота сердечных сокращений и пульсовое давление в arteria ascendens до полета и сразу же после взлета птицы. 50 мм по горизонтали соответствуют 1 сек, а 2,6 мм по вертикали - калибровочному давлению 250 мм вод. ст.
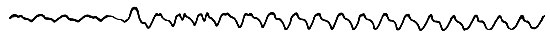
Фиг. 8. Частота сердечных сокращений и пульсовое давление морской чайки до полета и сразу же после взлета
Частота сердечных сокращений составляла 180 ударов в 1 мин до полета и 220 в 1 мин - сразу же после взлета. Таким образом, частота лишь слегка увеличилась, тогда как пульсовое давление, как явствует из кривых фиг. 8 и 9, возросло вдвое.

Фиг. 9. Кривая, полученная на другой морской чайке. Изменения диастолического давления не наблюдается
Это явление можно объяснить одним из двух способов. Можно предположить, что отмеченные гемодинамические сдвиги не имеют ничего общего с нормальной физиологией полета, а связаны с условиями опыта. Известно, что обратный ток крови из аорты в желудочек (регургитация) влияет на диастолическое давление в том же направлении (правда, в большей степени), что и снижение периферического сопротивления. Регургитация может быть следствием недостаточности аортальных клапанов, связанной с самим введением иглы или, что более вероятно, с ее смещением в процессе полета. Можно представить себе даже, что игла, сместившись, прокалывает один из аортальных клапанов и таким образом вызывает недостаточность. Мы пытались вводить иглу на разных расстояниях от клапанов, и оказалось, что даже^при введении ее в arteria brachicephalica, полностью исключающем ее соприкосновение с клапанами, результат не изменился.
Возможно, это явление объясняется электромеханическими возмущениями (при сильных движениях птицы на взлете), приводящими к изменению несущей частоты передатчика. Это более вероятно, так как в другом случае мы получили результат, приведенный на фиг. 9. Здесь нет заметного возмущающего воздействия, и диастолическое давление остается тем же, что и до полета.
Но вероятнее всего, что объяснение полученному результату следует искать в нормальной физиологии полета.
Если частота сердечных сокращений несколько повышается, а другие факторы остаются неизменными, то увеличение ударного объема крови приводит главным образом к повышению систолического давления. Диастолическое давление возрастает менее заметно. Следовательно, пульсовое давление увеличивается. Менее выраженное увеличение диастолического давления объясняется тем, что в результате высокого давления в конце периода изгнания становится круче градиент давления во время диастолы и большая часть энергии расходуется на скорость кровотока. Следовательно, большая (чем обычно) часть крови, выбрасываемой в артериальную систему во время систолы, успеет пройти через артериолы к концу диастолы.
Однако в опытах с домашней уткой пульсовое давление изменялось очень мало. В настоящее время мы не в состоянии дать какое-либо объяснение этим различиям в сосудистых реакциях на полет у разных птиц. Приведенные в работе данные для обеих птиц проверялись 15 раз, и результаты были одни и те же. Возможно, что они действительно отражают картину сердечно-сосудистых реакций на полет у плохо и у хорошо летающих птиц.
В заключение можно еще раз отметить, что, согласно полученным данным, частота сердечных сокращений в полете та же, что и до него, а пульсовое давление увеличивается.
Видеонаблюдение астрахань купить на сайте http://b2b.real.su.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'