Возможности использования телеметрии для записи биотоков мозга человека и животных, находящихся в космическом полете (У. Эйди)
Я очень коротко расскажу о работе, которую мы проделали за последние три года. Мы хотели проверить возможность записи электроэнцефалограмм (ЭЭГ) человека и животных во время космического полета (включая этапы разгона при подъеме и торможения при спуске). В этом докладе я коснусь четырех вопросов: 1) разработка методов вживления электродов для записи биотоков глубоко расположенных структур мозга; 2) разработка специальной аппаратуры для записи ЭЭГ и для тренировки животных; 3) специальная проверка на центрифуге и вибростенде при одновременной записи биотоков; 4) анализ полученных в полете данных с целью минимизации требований к телеметрической аппаратуре.
Мы проверяли методы вживления электродов в глубокие области мозга и часто сравнивали между собой различные типы электродов на одном и том же мозге. В одни участки мозга мы вводили жесткие электроды, представлявшие собой металлический стержень, из кончика которого была выведена тонкая проволочка. В симметрично расположенные участки мозга вводились тонкие проволочки без стержней. Записи, полученные с помощью этих электродов при проверке объекта на вибростенде и центрифуге, сравнивали между собой. Для определения повреждений мы исследовали эти участки мозга под микроскопом.
Участки мозга, наиболее чувствительные к изменениям физического и психического состояния, расположены в височной области. Исследования проводились на кошках, макаках и в последнее время - на шимпанзе. Мы вводили электроды в различные поверхностные и глубокие участки мозга, но большая часть моего сообщения относится к реакциям глубоких областей височных долей мозга. Часто спрашивают, возможно ли введение электродов в мозг без повреждения его при проверке объекта на вибростенде и центрифуге. Мы изучали влияние повторных испытаний на центрифуге (ускорения 8 g - 10 g) и на вибростенде. Через несколько месяцев после последнего испытания у таких животных не отмечалось более сильной реакции глии по ходу электрода, чем у животных, не проходивших проверок. Таким образом, довольно распространенное мнение о том, что при тряске электроды разрезают участки мозга, имеющего желеобразную консистенцию, не подтвердилось. Как бы то ни было, мозг не ведет себя подобным образом. Это мы проверяли много раз и в различных условиях.
На фиг. 1 изображена обезьяна (макак), привязанная к сидению центрифуги. К каждой из двух небольших пробок подходят провода от 18 электродов, которые связаны с усилительной системой при помощи специального кабеля, разработанного для управляемых снарядов и предназначенного для передачи очень малых сигналов. Конструкция кабеля позволяет минимизировать опасную статическую нагрузку на него. Между внутренней и внешней плетеными оболочками кабеля находится слой алюминиевой пудры, благодаря чему любые его изгибы не приводят к повреждениям проводов.

Фиг. 1. Обезьяна с электродами, введенными в глубоко расположенные участки мозга, привязанная к сиденью центрифуги. Записывались ЭЭГ обезьяны при решении ею задач по распознаванию образов, появлявшихся перед ней на специальной панели. Ясно видны крепления проводов к голове обезьяны
Подобным же образом мы произвели недавно вживление электродов в мозг шимпанзе. Трехлетний самец шимпанзе заранее был приучен к сиденью центрифуги. На его голове было укреплено обычное устройство с пробками. Этому предшествовала более чем годовая подготовительная работа. Мы использовали стереотаксический атлас, при помощи которого можно с точностью до 1 мм попасть в любую область мозга, зная вес тела и, конечно, размер головы. Это был, насколько мне известно, первый шимпанзе, которому ввели электроды и записали биотоки мозга.
Используемая аппаратура
Несколько лет назад мы разработали усилитель, удовлетворяющий нашим требованиям. Этот усилитель для снятия ЭЭГ очень устойчив и имеет коэффициент усиления около 40000. Все чувствительные к изменениям температуры элементы помещены в блок из сплава магния. После сборки и проверки усилитель помещается в оболочку из резины с эпоксидным наполнителем. Этот блок (при работе с максимальным коэффициентом усиления) выдержал испытания на вибростенде при частотах 2-5 кгц и ускорениях до 25 g. Усилитель имеет резонансный пик при 750 гц, но это не отражается на его рабочих характеристиках. Усилитель очень устойчив и способен выдержать любые внешние воздействия. Им можно даже забивать гвозди, и это не отражается на его нормальной работе.
На следующих этапах работы (сравнительно недавно) мы сконструировали микроминиатюрный усилитель, помещающийся в электроде, прикрепляемом к коже головы (фиг. 2 и 3). Эта конструкция предназначена в первую очередь для исследований человека в космическом полете. Трехкаскадный усилитель на транзисторах укреплен между двумя слоями нейлона. Размеры прибора - менее 12 мм по диаметру и по высоте. Он помещается в эпоксидную или силастиковую пластмассовую оболочку, к которой прикрепляется электрод-датчик, используемый для записи ЭЭГ. Сверху прибор окружает головка из нержавеющей стали, играющая роль экрана. Сигналы принимаются с его нижней стороны и поступают через слой губчатого вещества, пропитанного электродной пастой. Прикрепление к коже головы производится при помощи пробковой прокладки.

Фиг. 2. Микроминиатюрный трехкаскадный предусилитель на транзисторах, разработанный для записи ЭЭГ поверхностных участков мозга человека и животных. Все соединения выполнены при помощи пайки

Фиг. 3. Электрод вместе с усилителем, показанным на фиг. 2, в головке из нержавеющей стали. Нижняя прокладка обеспечивает надежное крепление к коже головы
Для специалистов по электронике могу сообщить, что входное сопротивление усилителя составляло 150 ком. Два таких прибора, включенные в дифференциальную схему, имеют входное сопротивление 300 ком. Выходное сопротивление прибора лишь 1500 ом, в связи с чем прибор очень устойчив к возмущениям, возникающим от сотрясения кабеля при повороте головы человека или животного. Коэффициент усиления прибора равен примерно 100, что обеспечивает выходные сигналы порядка нескольких милливольт. Эти сигналы могут поступать непосредственно на генераторы поднесущей частоты, используемые при телеизмерениях.
При опытах с центрифугой мы разработали специальную тренировочную панель. Подобная панель была приспособлена ранее на базе Военно-воздушных сил Холломан для опытов с шимпанзе. На трех небольших экранах, расположенных в передней части панели, одновременно появляются различные символы. Обезьяна узнает какой-то определенный символ, надавливает на него и получает в награду вкусную пилюлю. Отличие нашей панели от панели, использованной на базе Холломан, состоит в том, что у нас возможно несколько миллионов неповторяющихся комбинаций символов. Дело в том, что шимпанзе - очень высокоразвитая обезьяна, которая может быстро запомнить ограниченную последовательность символов, после чего она уделяет им очень мало внимания.
Для телеизмерений мы пользовались стандартным генератором поднесущей системы УИЗО (управляемый импульсами запросчик-ответчик). Это было удобно для нашей лабораторной работы, так как мы могли одновременно использовать стандартный прибор для регистрации данных на магнитной ленте. Запись велась на двух дорожках, причем на каждой из них регистрировались данные от 14 каналов телепередачи (фиг. 4). Хотя прибор накладывает некоторые ограничения на полосу частот и амплитуду сигналов, система в целом получается достаточно гибкой. Она имеет ценные преимущества при некоторых вычислениях, когда сигналы с головок, установленных на нескольких каналах, могут использоваться для задач, решаемых обычно при получении сигналов от различных записывающих устройств.

Фиг. 4. Сложная система с частотной модуляцией сигналов, предназначенная для записи биотоков мозга на магнитной ленте. Генераторы поднесущей и смесители расположены над устройством протяжки ленты. Система допускает одновременную запись 14 каналов на одной дорожке. Аппаратура очень надежна и великолепно отвечает всем требованиям эксперимента
Результаты испытаний
Для опытов с людьми и животными мы использовали большую центрифугу, развивающую ускорения до 10 g. С помощью этой центрифуги мы имитировали 14-дневный орбитальный полет обезьян в ракете "Атлас", в котором они самостоятельно питались на основе выработанного умения распознавать образы. В конце "полета" ускорения были близки к тем, которые наблюдаются при торможении ракеты.
В опытах с кошками при возрастании поперечного ускорения до 8 g наблюдалось резкое увеличение числа регулярно возникающих ритмических разрядов в височной области. Это явление исчезало, когда ускорение неизменно поддерживалось на величине 8 g. Когда ускорение начинает уменьшаться, оно возникает вновь, затухая при переходе центрифуги к вращению с постоянным ускорением. При таком увеличении продольных ускорений, когда животное теряет сознание, наблюдается весьма необычный разряд импульсов, напоминающий по форме эпилептический. Он возникает в глубоких областях височных долей мозга, распространяется на другие отделы и сопровождается движениями мышц и другими признаками эпилептического припадка. В некоторых случаях он не сопровождается двигательными реакциями, но, конечно, в это время полностью или в значительной степени пропадают все навыки животного по распознаванию образов.
В опытах с обезьянами при увеличении продольных ускорений наблюдались подобные же явления. Животное находилось в бессознательном состоянии, записываемые сигналы затухали. При резком уменьшении ускорения вновь появлялись судорожные разряды, после чего сознание восстанавливалось (фиг. 5).
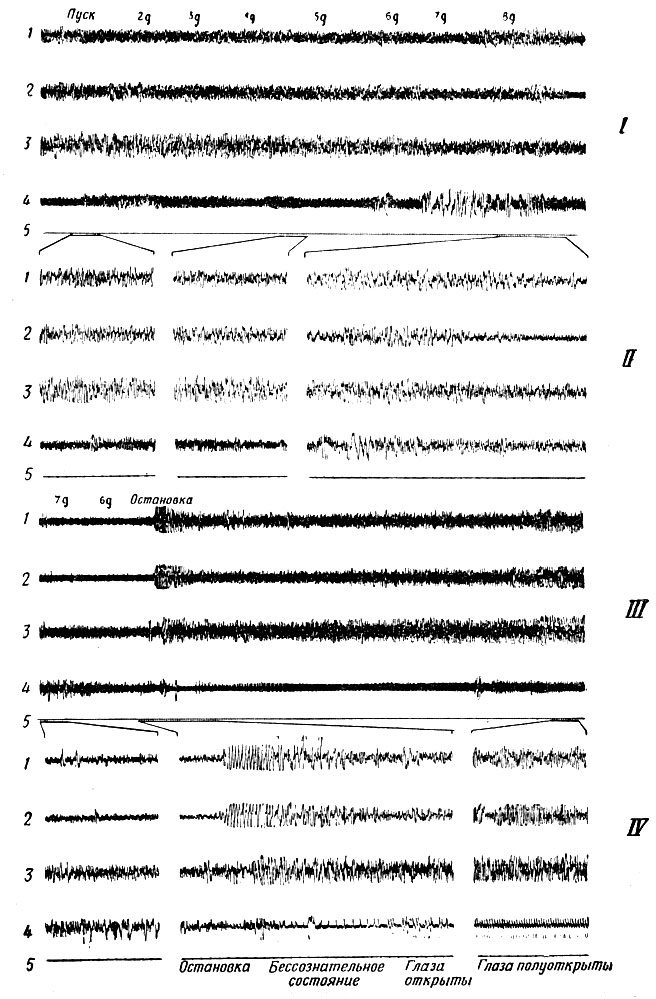
Фиг. 5. Образцы записей биотоков мозга (кривые 1-3 в каждой группе) и ЭКГ и ЭМГ (кривые 4) при испытании обезьяны на центрифуге до впадения в бессознательное состояние). 1 - левая миндалина (глубокая область височной доли); 2 - левая зрительная зона коры головного мозга; 3 - левый аммонов рог (глубокая область височной доли); 4 - электромиограмма и электрокардиограмма; 5 - отметка времени (1 сек). Группы записей I и III, выполненные при небольшой скорости протяжки ленты, показывают наступление бессознательного состояния и восстановление сознания. В группах II и IV приведены более подробные записи. Животное теряет сознание при ускорении 8 g (правый конец записей группы I). При остановке центрифуги (группа III) после серии электрических разрядов происходит восстановление прежнего вида ЭЭГ. Этот момент можно изучить более подробно по средней части записей группы IV. Записи получены Винтерсом, Кадо и Эйди, 1962 г
Два года назад мы начали исследование действия вибрации на обезьяну, одетую в костюм из пенопласта, в котором поддерживалось определенное давление. В последнее время мы используем иные методы, причем обезьяна прикрепляется к сиденью, напоминающему кресло космонавта.
При частоте вибрации около 10 гц появляются сдвиги ритмов электрической активности мозга. Когда мы заметили их впервые, то решили, что это просто артефакты. Но эти сдвиги наблюдались на определенных резонансных частотах и исчезали на других, причем они были разными для различных областей мозга и в разное время. Недавно мы обнаружили, что эти явления исчезают при гибели или усыплении животного. Ясно, что это не артефакты, а нечто, указывающее на существенную аномалию ритмов, вызванную вибрацией. Остаточных явлений после этого испытания мы не замечали. Животные, которых подвергали проверке на вибростенде 2 года назад (фиг. 6), находятся в отличном состоянии.
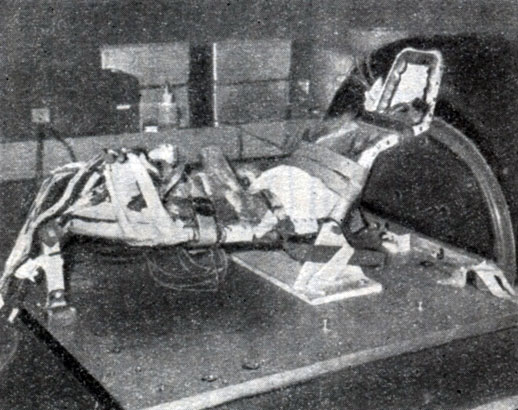
Фиг. 6. Обезьяна на вибростенде. Записывались биотоки из глубоких и поверхностных областей мозга при частотах вибрации 5-40 гц. При некоторых определенных частотах были заметны нарушения обычного вида ЭЭГ. Акселерометры на голове и теле обезьяны определяют величину ускорения
Для того чтобы выяснить, насколько целесообразной может оказаться запись ЭЭГ при длительном космическом полете для регистрации, скажем, циклов сон - бодрствование, мы непрерывно записывали сигналы из поверхностных и глубоких участков мозга у шимпанзе, находящегося в разных состояниях - от состояния бодрствования до наступления дремоты, сопровождающегося опусканием век и мочеиспусканием. В начале сна наблюдалось регулярное увеличение амплитуды волн. Не надо быть знатоком ЭЭГ, чтобы различить, спит или пробуждается животное, так как регулярное возникновение волн с большой амплитудой дает, как мы называем, "игольчатую" запись. Когда животное просыпается, садится и смотрит вокруг, ЭЭГ приобретает совершенно иной характер (фиг. 7).
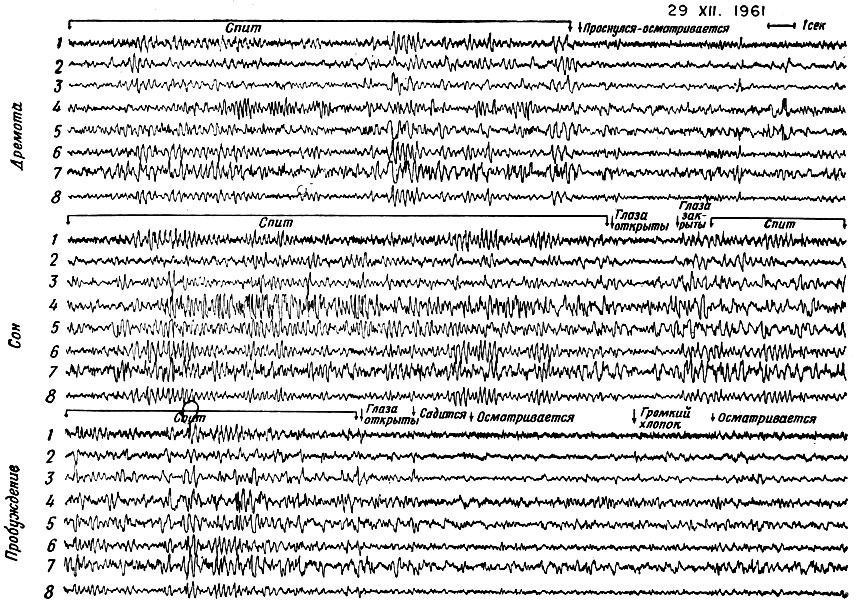
Фиг. 7. Записи биотоков из поверхностных и глубоких областей мозга шимпанзе. 1 - левая миндалина; 2 - правая миндалина; 3 - передняя часть левого аммонова рога; 4 - лобно-теменная зона коры (поверхностное отведение); 5 - левая обонятельная зона коры (глубокая область височной доли); 6 - задняя часть правого аммонова рога; 7 - ретикулярная формация среднего мозга (центральная часть нижнего отдела ствола мозга), слева; 8 - затылочно-теменная зона коры (поверхностное отведение). Впервые в подобных исследованиях шимпанзе были введены в мозг электроды для записи ЭЭГ. На записях видно начало регулярных колебаний большой амплитуды, когда внимание уменьшается, наступает дремота и сон. Записи при усталости и сне явно отличны от более высокочастотных, небольших по амплитуде и менее регулярных ЭЭГ бодрствующего животного
Итак, можно с уверенностью сказать, что имеется уже одна область, в которой ЭЭГ может дать денную информацию. Есть основания думать, что при длительной невесомости возможно нарушение циклов сон - бодрствование. Известно, что дельфины спят очень мало. Вероятно, что так же мало спал бы человек, если бы он находился в состоянии невесомости без одежды и при температурном равновесии с окружающей средой. Нам кажется, что все это стоит дальнейшего изучения.
Методы уменьшения избыточности информации
Наконец, мы переходим к вопросу об уменьшении избыточности информации, содержащейся в данных, полученных при помощи телеизмерений. Если бы мы смогли устранить необходимость телепередачи большого количества необработанных данных, то это было бы весьма выгодно как для самого эксперимента, так и для уменьшения ошибок в полученных данных. В одной из наших лабораторий мы работаем над выбором наиболее подходящей формы для записи данных и над методом их обработки с целью уменьшения избыточности. Информация от приборов, находящихся в этой лаборатории, может поступать непосредственно на вход большой вычислительной машины IBM 7090. Я хотел бы охарактеризовать тип задач, которые можно решать с помощью таких машин, так как это помогает понять важность совместной работы биологов и инженеров. Я неизлечим от желания "делать все самому", но тем не менее должен сказать, что именно та информация, которую мы можем получить из записей биотоков мозга, полученных при решении животным задачи распознавания, ясно показывает, насколько важен наш союз с инженерами.
В типичных ЭЭГ животного, записанных при решении им задачи распознавания символа (для получения пищи), по характеру волн мозга четко различаются период "перед распознаванием" и период "распознавания". Все наши ЭЭГ, кроме одной, записаны из глубоких участков мозга, и лишь одна запись сделана из зрительной зоны коры. Для того чтобы по возможности просто проанализировать наши записи и установить природу этих волновых процессов, мы воспользовались методом автокорреляционного анализа. Вычисления показали существенное различие между ЭЭГ в период "перед распознаванием" и в период "распознавания". Мы были поражены, обнаружив, что на основе этого анализа мы можем отличить правильные действия животного от неправильных.
Например, можно построить взаимно-корреляционную функцию фазы колебаний в различных участках мозга. Я не буду останавливаться здесь на том, как мы измеряли фазу. Для правильных реакций животного эта корреляционная функция имеет один вид, а для неправильных - другой. Мы сравнивали результаты по различным дням обучения и обнаружили, что взаимно-корреляционные функции совпадают для всех случаев правильных реакций животного, а также для всех неправильных реакций, причем между первыми и последними имеется существенное отличие. Это очень интересное применение метода, хорошо известного математикам и инженерам, использующим его для решения проблем вибрации управляемых снарядов и т. п. Теперь этот метод занял определенное место в анализе ЭЭГ.
Мы использовали значительно более сложный метод анализа, который позволил нам определить взаимно-корреляционные функции по амплитуде и фазе во всем спектре частот. Этот метод анализа, который также был разработан для исследования вибраций ракет, позволяет, например, определить корреляцию фазы колебаний в двух участках мозга на частотах от 2 до 20 гц. Мы обнаружили, что при правильной реакции животного эти колебания на частотах 2-12 гц сдвинуты по фазе на +30°. При неправильной реакции животного в фазовом угле происходят существенные изменения, причем на частоте 5 гц происходит сдвиг фазы с +90° до -90°. Мы наблюдали подобные явления у разных животных и в различных ситуациях, и нас просто поражало постоянство различия в сдвиге фаз при неправильных и правильных реакциях.
Мы использовали также математический метод, разработанный для анализа показаний магнитометра, находящегося на спутнике Земли. Изменения магнитного поля Земли производят очень малую фазовую модуляцию синусоидального выходного сигнала. Если бы магнитное поле Земли было постоянным, то никакой модуляции не происходило бы и на выходе прибора была бы идеальная синусоида.
Мы использовали этот метод для анализа ЭЭГ, полученных при распознавании животным определенного символа (для получения пищи). Записанные при этом колебания имеют почти стабильную частоту. Полученный результат можно представить как модуляцию некоторой "центральной частоты" порядка 5,5 гц. Эта частота появляется только в моменты наибольшего внимания животного.
Я знаю, что в цели настоящей конференции не входит подробное обсуждение вопроса об обработке полученных данных, и лишь подчеркну, что подобные методы обработки позволяют получить очень ценную информацию из весьма сложных записей.
Обсуждение доклада
Корсон. Одинаково ли влияет вибрация на электрическую активность всех участков мозга? Нельзя ли исследовать эту активность, применяя некоторые успокаивающие средства, например фенотиазин или какие-либо препараты, расслабляющие мышцы?
Эйди. В первую очередь нас интересуют источники тех резко выраженных изменений, которые мы наблюдаем в ЭЭГ. Их, по-видимому, следует искать в двух системах: мышечно-суставной и вестибулярной.
К областям мозга, максимально реагирующим на внешние возмущения, относятся ретикулярная система ствола мозга, некоторые первичные чувствительные области и височная доля. В этих областях ритмы, наблюдаемые при решении животным какой-либо задачи с целью получения пищи, наиболее заметны. Зрительная зона коры действует даже тогда, когда глаза животного завязаны. Для исключения фактора зрительной стимуляции мы проводили несколько опытов с обезьянами, у которых были завязаны глаза. Таким образом, имеются некоторые определенные области мозга, проявляющие наибольшую чувствительность. Однако о путях возникновения этой активности мы ничего не знаем.
Дельгадо. Не ясно, что было признано лучшим: стержневые или гибкие электроды? Второй вопрос: в какой области мозга наблюдается наиболее медленное восстановление ритма по окончании действия ускорения? Я думаю, что, говоря о височной доле, вы в основном имеете в виду аммонов рог.
Эйди. Очень интересно, что именно аммонов рог особенно медленно приходит в нормальное состояние после действия длительных ускорений. На это уходит от 30 сек до 1 мин, а для миндалины - еще больше - 2-3 мин.
Относительно качества электродов я могу сказать, что, тщательно изучив множество ЭЭГ, зарегистрированных при испытаниях на центрифуге, мы не нашли различия в кривых, полученных с помощью гибких и с помощью стержневых электродов.
При гистологическом изучении было обнаружено, что стержневые электроды не вызывали серьезных повреждений в месте их ввода в мозг. Мы не обнаружили существенных различий между глубокими (ниже 10-15 мм) участками мозга животных, испытывавшихся на центрифуге, и других животных, не проходивших эти испытания. Возможно, что при ускорениях, создававшихся в наших опытах, мозг ведет себя как вязкая жидкость, в котором перемещаются лишь самые поверхностные слои. Я не знаю, так ли все это, но мы считаем такое предположение правильным.
Маккей. Были ли расположены электроды перпендикулярно к направлению ускорения?
Эйди. Нет, они были расположены произвольно.
Мак-Каллок. Каков был метод вычисления корреляционных функций сдвига по фазе?
Эйди. Мы применяли метод взаимно-корреляционного анализа. Первоначально использовался механический коррелятор. Затем мы производили запись на магнитной ленте и использовали магнитный коррелятор. В последнее время мы используем вычислительную машину с очень большой программой, составленной для исследования вибраций ракет. Мы получаем авто- и взаимно-корреляционные функции и соответствующие спектральные плотности.
Интересен следующий факт, хотя он, возможно, расстраивает наши планы. Если взять записи 4 каналов ЭЭГ на отрезке времени 200 сек и прерывать запись каждого канала 167 раз в 1 сек, то мы получим сравнительно небольшое количество данных. Однако вычислительной машине IBM7090 при соответствующей программе нужно 90 мин для исследования этих данных. Это не является, строго говоря, уплотнением данных.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'