Дыхание микробов
Дыхание является источником энергии для живых существ. Происходящие в микробной клетке синтетические процессы построения протоплазмы, процессы роста, размножения, движения и др. требуют притока свободной энергии, так как эти процессы эндотермичны. Поэтому в микробной клетке постоянно совершаются одновременно с процессами ассимиляции процессы диссимиляции, освобождающие энергию для ее жизнедеятельности.
Совокупность биохимических процессов, в результате которых образуется энергия, необходимая для жизнедеятельности клетки, составляет энергетический обмен. В противоположность высшим организмам энергетический обмен у микроорганизмов имеет разнообразные формы: дыхание, брожение и др.
Дыханием называется окисление органических веществ с помощью газообразного кислорода до углекислоты и воды. Так, окисление сахара в процессе дыхания выражается уравнением С6Н12О6+6О2=6СО2+6Н2О+энергия. Это уравнение противоположно уравнению фотосинтеза 6СО2+6Н2О+энергия=C6H12О6+6O2.
В 1861 г. Луи Пастер при изучении масляно-кислого брожения установил, что возбудитель этого брожения (Вас. butyricus) нормально развивается только в отсутствие свободного кислорода, энергию он поручает за счет реакции расщепления органического субстрата. Пастер определил сущность брожения как жизнь без кислорода. Кислород воздуха не принимает участия в брожении, а органическое вещество окисляется за счет отнятия водорода, который присоединяется к продуктам распада этого же органического вещества или выделяется в газообразном состоянии.
Автотрофы получают энергию за счет окисления простых неорганических соединений: сероводорода, аммиака, водорода. Денитрифицирующие и десульфофицирующие бактерии получают энергию путем окисления соответственно нитратов и сульфатов, но они могут получать энергию и за счет окисления органических веществ. У некоторых микробов реакция окисления кислородом не доходит до конечных продуктов - СО2 и Н2О. Такой неполный окислительный процесс наблюдается у уксуснокислых бактерий, окисляющих спирт только до уксусной кислоты, и некоторых видов плесеней, разлагающих сахар до щавелевой и лимонной кислот.
Гнилостные бактерии используют энергию, освобождающуюся при расщеплении ими белков, при этом энергия химических связей аминокислот превращается в энергию АТФ.
По отношению к кислороду микробы разделяются на две группы: аэробы, развивающиеся только при наличии кислорода в окружающей среде, и анаэробы, которые развиваются при отсутствии свободного кислорода. Кроме того, имеется еще одна промежуточная группа - факультативные анаэробы, способные жить как в аэробных, так и в анаэробных условиях. Имеются также микроаэрофилы, развивающиеся при уменьшенном количестве кислорода в среде, например бруцеллезная палочка.
Образующаяся химическая энергия только частично рассеивается в виде тепла. Большая же часть этой энергии улавливается и сохраняется в виде макроэргических связей АТФ. Фосфатные группы связаны между собой непрочно и легко освобождают свою энергию в нужных количествах там, где она необходима для жизнедеятельности клетки. АТФ, теряя энергию, превращается в АДФ (аденозиндифосфат) и в АМФ (аденозинмонофосфат).
АТФ, АДФ, АМФ и фосфорная кислота всегда присутствуют в различных соотношениях в каждой клетке. Реакции эти обратимы, АМФ и АДФ могут присоединять к себе фосфорную кислоту и превращаться в АТФ. Так обеспечивается более или менее постоянное количество АТФ в клетке. Запас АТФ в клетке ограничен. Для восстановления макроэргических связей АТФ все время используется энергия расщепления углеводов и других веществ.
Долгое время считали, что процесс дыхания свойствен высшим организмам, а брожение только микроорганизмам. Затем было установлено, что они тесно связаны друг с другом. Дыхание и брожение представляют собой очень сложные комплексы сопряженных окислительно-восстановительных процессов, которые определяются тем или иным набором ферментов.
Во всех энергетических процессах в клетке можно различить три стадии. В первой, подготовительной стадии крупные молекулы углеводов, жиров, белков распадаются на небольшие молекулы глюкозы, глицерина, жирных кислот, аминокислот. Происходит подготовка веществ к дальнейшим превращениям, здесь не происходит заметного извлечения энергии.
В следующей стадии, называемой стадией неполного окисления, образовавшиеся глюкоза, жирные кислоты и другие вещества подвергаются сложному многоступенчатому процессу. Таково неполное окисление, которое называется гликолизом или брожением. Эта стадия анаэробная. Гликолиз представляет собой более десяти последовательных ферментативных реакций. Из глюкозы последовательно образуется десять различных промежуточных веществ (субстратов) и действует столько же специфических ферментов. Весь этот процесс идет по типу молочнокислого брожения, вызываемого молочнокислыми бактериями, и имеет много сходного со спиртовым брожением, вызываемым дрожжами.
Процесс начинает фермент гексокиназа, под влиянием которого глюкоза вступает в реакцию с АМФ и образуется гексозо-6-фосфат. Гексозо-6-фосфат под действием фермента алдолазы переходит во фруктозо-6-фосфат и т. д. Конечным продуктом гликолиза является молочная кислота. Суммарное уравнение всего процесса выражается так: С6Н12О6=2С3H6О3.
Последней стадией процесса является полное окисление субстратов до конечных продуктов - СО2 и Н2О. Эта стадия протекает в аэробных условиях. Следовательно, она имеется только у аэробов. В этой стадии участвуют органические кислоты, состоящие из трех углеродных атомов, почему и назвали ее циклом трикарбоновых кислот (Крепс, 1953).
Цикл начинается с того, что две молекулы молочной кислоты окисляются и дают две молекулы пировиноградной кислоты. Одна из молекул пировиноградной кислоты окисляется с отщеплением одной молекулы углекислоты, и образуется уксусная кислота. Углекислота связывается с другой молекулой пировиноградной кислоты, и образуется щавелевоуксусная кислота. Уксусная кислота соединяется с коферментом А, происходит конденсация ее с щавелевоуксусной кислотой и водой, и образуется лимонная кислота.
Лимонная кислота превращается в аконитовую кислоту. Далее происходит еще ряд превращений с образованием снова щавелевоуксусной кислоты, и па этом цикл заканчивается. Лимонная кислота оказывается вся разложенной. Из ферментов в цикле участвуют дегидрогеназы НАД, ФАД, цитохромы. Так, при дегидровании янтарной кислоты в цикле от нее отнимается электрон, он переносится на ФАД, и образуется ФАД-Н2, янтарная кислота окисляется в фумаровую кислоту. Электроны далее продвигаются по цепочке цитохромов к кислороду. Происходит соединение с активированным цитохромоксидазой кислородом с образованием воды. Непосредственно же кислород в реакциях не участвует.
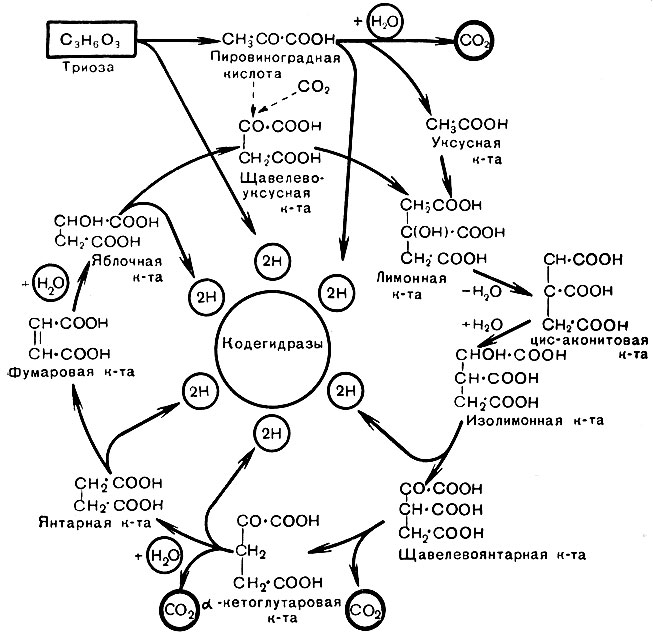
Рис. 19. Схема цикла трикарбоновых кислот (цикл Кребса)
Подсчитано, что в этом цикле из одной молекулы глюкозы образуется 36 молекул АТФ и в процессе гликолиза две молекулы АТФ, всего, следовательно, 38 молекул АТФ, или 380 больших калорий из 680 больших калорий, содержащихся в грамм-молекуле глюкозы, т. е. получено 55% полезной химической энергии. Это очень большой процент но сравнению с к.п.д., получаемым в технике (12-25%). Энергия выделялась постепенно по частям. Если бы она была выделена сразу, то клетка была бы повреждена.
Ферменты, принимающие участие в реакциях, расположены в митохондриях и локализованы рядами по порядку их действия в ходе гликолиза и цикла трикарбоновых кислот.
Продукты распада триоз частично идут на биосинтез. Так, из пировиноградной кислоты может образоваться аланин, из кетоглутаровой кислоты - глютаминовая кислота, из щавелевоуксусной кислоты - аспарагиновая кислота путем аминирования. Уксусная кислота может пойти на синтез высших жирных кислот.
Протекающие при дыхании реакции имеют сопряженный характер окисления-восстановления. В ходе реакций окисления-восстановления развивается электродвижущая сила, которая может быть измерена в виде так называемого окислительно-восстановительного потенциала (rН2).
Аэробы приспособлены к более высокому rН2 (20 и выше), анаэробы - к низкому (0-12), факультативные анаэробы - 0-20. Снижая rН2 среды, можно добиться роста анаэробов в присутствии кислорода, а повышая rН2, выращивать аэробы в анаэробных условиях.
В энергетическом отношении анаэробное дыхание во много раз менее эффективно, чем аэробное. Так, если при аэробном процессе окисления глюкозы до СО2 и Н2О получается 674 ккал, то при спиртовом брожении - 27 ккал, при молочнокислом - 18 ккал и при масляно-кислом - всего 15 ккал. Это объясняется тем, что конечными продуктами анаэробного окисления являются органические соединения, сохранившие еще большой запас энергии. Например, спирт (продукт спиртового брожения) прекрасно горит.
Потерю тепла при брожении можно наблюдать в культурах, хорошо защищенных от потери тепла. За счет выделения этого тепла происходит самонагревание влажного сена, навоза, торфа и пр.
У светящихся бактерий потеря энергии выражается в виде свечения. Свечение морской воды, гнилого дерева, мха, рыбы объясняется присутствием на них особых светящихся бактерий аэробов. Они имеют особый фермент - люцеферазу, который химическую энергию АТФ переводит в энергию света.
К анаэробным относятся палочки столбняка, ботулинуса, масляно-кислые бактерии, возбудители газовой гангрены и др.
К аэробам относятся нитрифицирующие, уксуснокислые, азотобактерии, миксобактерии, плесени, микобактерии, холерный вибрион. Факультативные анаэробы - кишечная палочка, дифтерийная палочка, стрептококк, стафилококк, спириллы и др.
Порнуха. Новое порно: Ru Soska. Онлайн порно.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'