Глава 7. Общая микробиология. (В. Н. Гутина)
Особенности микробиологии в XX веке
Создание широких теоретических обобщений и успешное практическое применение результатов научных исследований составили основное содержание прогресса этой отрасли знания в XX в. Характерной чертой развития микробиологии в XX в. является все более углубляющаяся дифференциация, связанная с открытием и изучением большого разнообразия микроорганизмов и приведшая к обособлению новых научных дисциплин - бактериологии, микологии, вирусологии. Кроме дифференциации по систематическому признаку в микробиологии выделились научные разделы, различающиеся объектами и задачами исследования. Таковы общая, медицинская, сельскохозяйственная, почвенная, техническая, водная, геологическая, радиационная микробиология и генетика микроорганизмов. Из них общая микробиология, изучающая строение и жизнедеятельность всех видов микроорганизмов, приобрела интегрирующее значение.
Опережение разработки теоретических представлений о механизме микробиологических процессов их практическим использованием характерно не только для раннего, но до некоторой степени и для современного этапа развития микробиологии, когда результаты практического использования микробов служат ключом к выяснению их физиологических особенностей. Указанная методологическая закономерность развития микробиологии определяется спецификой самого объекта исследований.
В настоящее время микроорганизмы наряду с другими микроскопическими организмами служат удобными объектами генетических, биохимических, цитологических, физиологических и других исследований, на которых изучаются актуальные проблемы соответствующих наук.
Выявление большого разнообразия мира микробов повлекло за собой изучение разнообразия их химической активности, первые общие представления о которой давала практика. Раскрытие в первой половине XX в. таких форм биохимической деятельности микробов, как разложение, синтез и трансформация веществ субстрата, определило интенсивное развитие физиолого-биохимического направления в микробиологии, в результате чего был накоплен большой фактический материал по физиологии и биохимии микроорганизмов.
Наряду с обнаружением огромного многообразия физиологических функций микроорганизмов одной из ведущих теоретических проблем общей микробиологии в XX в. стало выявление единства живого путем изучения общих генетических и биохимических механизмов.
Широкое практическое использование разнообразных форм жизнедеятельности микробов сделало в XX в. актуальной теоретическую и практическую разработку вопросов культивирования микроорганизмов с целью интенсификации вызываемых ими процессов. Это в свою очередь обусловило необходимость изучения основ регуляции роста и развития микроорганизмов, поиск способов воздействия на их обмен веществ, что определило формирование еще одного направления современной микробиологии - управляемого культивирования микроорганизмов.
Успехи почвенной, сельскохозяйственной, геологической, медицинской микробиологии в XX в. могут быть предметом специального исследования. В настоящей главе кратко представлено развитие таких направлений общей микробиологии, как изучение экологического разнообразия микробов, принципы их систематики и классификации, морфология и цитология микроорганизмов, а также изучение некоторых форм диссимиляционной, биосинтетической и трансформирующей деятельности микробов. Кроме того, предпринята попытка осветить основные результаты в решении проблемы управляемого культивирования и в развитии генетики микроорганизмов.
Выявление экологического разнообразия мира микробов
Огромное разнообразие микроорганизмов стало известно уже в конце; XIX в. В XX в. оно получило всестороннее экспериментальное обоснование в результате эколого-физиологических исследований. Выявление разнообразия форм жизнедеятельности микроорганизмов сопровождалось поисками в природе различных экологических типов. Экологическая микробиология оказалась в XX в. одной из основных сфер развития идей В. И. Вернадского (1923) о ведущей роли живых существ в превращениях химических элементов на земной поверхности. Трудами С. Н. Виноградского, В. Л. Омелянского, М. Бейеринка и других была доказана важнейшая роль микроорганизмов в геохимических процессах и установлено их огромное геологическое значение.
В первой половине XX в. экологическое направление в микробиологии особенно плодотворно развивалось в нашей стране. Идея о важнейшей роли микроорганизмов в круговороте веществ в природе была впервые высказана В. Л. Омелянским (1909). Он же представил схемы круговорота веществ в природе, указав, что путь к познанию закономерностей круговорота состоит в изучении морфологии, физиологии и экологии микроорганизмов. Омелянский провел детальное изучение жизнедеятельности микроорганизмов, принадлежащих к различным эколого-физиологическим группам: участвующих в круговороте азота - нитрификаторов и азотфиксаторов; осуществляющих круговорот серы - гнилостных и сульфатредуцирующих, бесцветных, окрашенных и тионовых бактерий; играющих важную роль в круговороте железа - железобактерий; обеспечивающих круговорот углерода - разложение и синтез органических соединений.
Успехи, достигнутые в раскрытии экологических особенностей микробов, были в значительной мере связаны с применением специальных методов изучения живой микрофлоры, основывающихся на принципе культивирования в среде, максимально приближающейся к природным условиям обитания микробов. Культивирование в таких особых условиях, не вызывающих нарушений в нормальной жизнедеятельности микробов, позволило воспроизвести многие микробиологические процессы и дало возможность контроля за развитием культуры. Экологическое направление в микробиологии обеспечило интенсивное развитие многих разделов этой науки. Идея С. Н. Виноградского (1924) о том, что только продолжительное по мере возможности наблюдение над процессом развития единичных форм (особей) в микрокультурах может привести к цели, легла в основу разработки методов прямого микроскопического исследования микроорганизмов.

Сергей Николаевич Виноградский (1856 - 1953)
Среди них особенно широкое применение получил метод стекол обрастания Росси - Холодного (1927, 1930). В применении к илам этот метод был усовершенствован гидробиологом Э. Науманом (1923). К числу подобных же приемов изучения микрофлоры в естественных условиях относятся: "почвенная камера" Н. Г. Холодного (1933) и его же метод "проращивания" почвенной пыли, представляющий собой модификацию метода почвенных комочков, созданного С. Н. Виноградским; приемы изучения глубоководной (баротолерантной) микрофлоры, разработанные А. Е. Криссом (1959; Ленинская премия, 1960) и его сотрудниками; капиллярные методы изучения микроорганизмов, введенные Б. В. Перфильевым и Д. Р. Габе (1961; Ленинская премия, 1964); пластинки из нитратного агара, предложенные Е. 3. Теппер для изучения проактиномицетов; педоскопы, созданные Т. В. Аристовской для изучения почкующихся бактерий и нашедшие широкое применение в почвенной микробиологии, и др.
В последние годы микробиологические исследования вод, илов, подводных грунтов, почв и горных пород, ставящие целью изучение состава свободно живущей микрофлоры и экологических закономерностей ее распределения, развивались менее интенсивно, чем до 60-х годов. Это связано с переключением внимания микробиологов на физиолого-биохимические, цитологические и генетические проблемы.
Развитие принципов систематики микробов
Известно, что состояние систематики организмов свидетельствует об уровне их познания и отражает преобладающие тенденции в развитии науки. Это правило наглядно подтверждается на примере развития микробиологии.
В XX в. классификация микробов развивалась на основе уже имевшихся систем микроорганизмов. Первые системы строились по принципам ботанической классификации и опирались преимущественно на морфологические признаки. Идея о необходимости использования при систематизации бактерий физиологических признаков была обоснована в трудах С. Н. Виноградского и М. Бейеринка и впервые реализована в системе молочнокислых бактерий С. Орла-Иенсеном (1919). Последующие попытки систематизации бактерий основывались преимущественно на принципе номенклатурных типов (Р. Бьюкенен, 1926), что привело к созданию определителей, носивших преимущественно характер описаний отдельных родов.
В первой половине XX в. в принципах классификации наметились два основных подхода.
В основе первого из них - филогенетического - лежит дарвиновская концепция эволюции и идея создания общей схемы происхождения и родства организмов. Методологической основой попыток построить генеалогическую систему микроорганизмов служил тезис о том, что источником информации об эволюции и филогенетическом родстве микроорганизмов могут быть результаты изучения современных форм. Правомерность этой точки зрения была в значительной мере подкреплена исследованиями генетиков и биохимиков, создавших основы геносистематики, которая дала возможность определять сходство и различие организмов в естественных единицах информации (А. Н. Белозерский, А. С. Спирин, Б. Ф. Ванюшин, Д. Де-Лей, Л. Хилл, Дж. Мармур и др.). Ведущим методом этого направления стало определение состава и последовательности расположения нуклеотидов в ДНК. С его помощью удалось в значительной мере пополнить представления о филогенетических связях в мире микроорганизмов. В то же время выяснилось, что при определении критериев родства этот метод может играть лишь вспомогательную роль.
Другой подход к классификации микробов отрицает значение их филогенетических и генеалогических особенностей. К сторонникам этой точки зрения относятся главным образом зарубежные исследователи (Р. Бьюкенен, Д. Жилмур, П. Снет, А. Коэн, С. Коэн и др.). Не отвергая наличия филетических связей между организмами, они считают, что на современном уровне знаний между ними если и можно установить известную степень родства, то лишь относительную. При этом основное таксономическое значение придается признакам, которые подлежат чисто статистической обработке (нумерическая таксономия). Все такие признаки рассматриваются как равноценные. В бактериологию этот метод был введен П. Снетом в 1957 г.
Следует отметить, однако, что в систематику бактерий принцип нумерической таксономии не внес ничего принципиально нового; он позволил лишь сравнительно быстро систематизировать большое многообразие форм.
Теоретические основы принятой в настоящее время систематики были разработаны А. Клюйвером и К. Ван-Нилем (1936), которые рассматривали род бактерий в качестве морфофизиологического единства. Вслед за этими исследователями стали считать, что в систематике бактерий, в отличие от систематики растений и животных, центральное положение занимает род, а не вид. При бедности морфологических типов организации и разнообразии физиологических функций бактерий некоторые исследователи (Р. Стениер, 1968) отдают предпочтение в таксономии физиологическим признакам. До сего времени нет общепринятого списка признаков-дескриптеров, который позволил бы унифицировать описание родов у прокариотов и вместе с данными геносистематики и филогенетики создал бы основу естественной систематики микроорганизмов.

Мартин Бейеринк (1851 - 1931)
Морфология и цитология микроорганизмов
Морфо-цитологические исследования микроорганизмов в XX в. развивались в направлении изучения ряда специальных проблем, сопровождавшегося разработкой как общих морфо-цитологических методов исследования (например, цитологических и цитохимических методов, совершенствованием светооптической и электронно-микроскопической техники), так и созданием некоторых специальных методов, обеспечивающих наблюдение за живыми, активно функционирующими клетками микробов. С помощью методов, разработанных Н. Г. Холодным, А. С. Разумовым, В. Д. Тимако-вым, М. А. Пешковым, Б. В. Перфильевым и другими, была выявлена в 20-х годах ошибочность так называемой теории циклогении Ф. Лениса, Г. Эндерлейна и Е. Алмквиста, утверждавшей наличие у микроорганизмов сложных циклов развития, протекающих по замкнутому кругу.
Начиная с 40-х годов морфология микроорганизмов стала развиваться преимущественно в направлении изучения структуры и функций клеточных органоидов. В нашей стране это направление получило развитие в трудах М. Н. Мейселя и его сотрудников.
Основными вопросами морфо-цитологических исследований микроорганизмов в XX в. были структура и химический состав ядерного аппарата и цитоплазмы, клеточной стенки, мембранной системы, митохондрий, рибосом, жгутиков и т. д. Эти исследования содействовали выявлению к концу 50-х годов цитологических различий между прокариотами и эвкариотами.

Альберт Клюйвер (1888 - 1956)
Данные о тинкториальных свойствах клеточной оболочки бактерий начали появляться с 1924 г. (М. Гутштейн, Г. Найзи, Р. Маррей, М. А. Пешков и др.). Было установлено, что известное ранее характерное окрашивание бактерий по Граму обусловлено содержанием, глубиной залегания и степенью защищенности липидами мукопептидного комплекса, ответственного за ригидность клеточной стенки.
Для изучения жгутиков с начала XX в. стали использоваться методы темного поля (К. Рейхерт, Ф. Науман), а позднее и фазово-контрастной микроскопии (А. Флеминг). А. Пийпер рассматривал жгутики как пассивные выросты клеточной стенки полисахаридной природы. Согласно другой, оказавшейся более правильной точке зрения, жгутики представляют собой органоиды, содержащие сократительные белки (Ч. Вейбулл). По данным Ф. Фурмана (1910), жгутики берут начало в базальной "грануле" (мембранная структура), расположенной в цитоплазме.
В конце 50-х - начале 60-х годов было показано, что цитоплазматическая мембрана у бактерий инвагинирует в цитоплазму, образуя внутриклеточные мембранные структуры - мезосомы (П. Фиц-Жаме, 1960) и хроматофоры. Оказалось, что эти структуры, количество которых в клетке может быть различным, обладают различной ферментативной активностью. К настоящему времени мнения о их роли в клетке разделились. Одни считают, что мезосомы - структуры полифункциональные, другие относят их к функционально строго дифференцированным образованиям. Предполагается, что мезосомы содержат ферменты дыхательной цепи (М. Солтон, В. М. Кушнарев и др.), ферменты азотфиксации (В. А. Яковлев с сотрудниками), участвуют в спорообразовании (П. Фиц-Жаме, Д. Охайе, У. Мурелл) и клеточном делении.
Интенсивно изучалось бактериальное ядро. Введение в практику цитологических исследований основных красителей позволило А. Мейеру в 1908 г. сформулировать положение о дискретности ядерной субстанции у бактерий. Согласно другой точке зрения, хроматин в бактериях диффузно распределен в цитоплазме и лишь при различных патологических состояниях или на ранних стадиях спорообразования "отмешивается" в виде гранул и тяжей (Ф. Шаудин, Р. Гертвиг, К. Добелл, К. Пиетшманн, А. А. Имшенецкий).
Применение реакции Фёльгена-Россенбека, контрастной фотографии, фазово-контрастного устройства, электронно-микроскопических, генетических и биохимических методов исследования привело в настоящее время к установлению того, что ядро, или нуклеоид, бактерий представлен в виде одной нити ДНК, свернутой в кольцо и прикрепленной к мембране, и что нуклеоид, или бактериальная хромосома, не имеет ядерной мембраны. В этом его главное отличие от структуры ядерного аппарата эвкариотов.
Богатый материал был получен по изучению функциональной морфологии органелл низших эвкариотов. Оказалось, что митохондрии дрожжей обладают большой изменчивостью структуры, высокой сорбционной активностью и легкой повреждаемостью при химических и физических воздействиях. Это подтвердило их участие в метаболических процессах. В 1938 г. М. Н. Мейсель описал характерную перестройку митохондрий у факультативных аэробных дрожжевых организмов при их переходе от аэробных к анаэробным условиям. Та же перестройка была обнаружена в 50-х годах при изменении концентрации Сахаров, витаминов и антибиотиков. Изучение ультраструктуры дрожжевых митохондрий выявило наличие у них крист с грибовидными субъединицами. Было показано, что эти же ультраструктуры характерны для всех самостоятельно существующих организмов, начиная от бактерий и кончая клетками высших организмов.
В 40 - 50-х годах было установлено, что внутренняя мембранная система, образующаяся из цитоплазматической мембраны, имеется главным образом у грамположительных бактерий; у грамотрицательных бактерий она развита слабее. Возникла гипотеза о гомологичности органелл эвкариотных клеток с целой клеткой прокариот.
Последние пять - семь лет ознаменовались значительным прогрессом в изучении морфогенеза бактерий и раскрытии его химической основы. Это стало возможным благодаря исследованиям генетического аппарата и биосинтеза структурных компонентов клетки.
Физиология и биохимия микроорганизмов
В формировании физиолого-биохимического подхода к изучению жизнедеятельности микробов ярко проявилась одна из характерных черт микробиологии, о которой говорилось выше: исследования, носящие прикладной характер, оказались источником данных, имеющих большую теоретическую и научную ценность. Вместе с тем специфические свойства микроорганизмов - быстрый рост и высокая активность обмена, разнообразие форм анаболизма и катаболизма, экологическая специфика - определили исключительную роль микроорганизмов как наиболее удобных объектов для химического изучения обмена веществ.
В теоретическом истолковании результатов физиолого-биохимических исследований выявились две взаимосвязанные тенденции: с одной стороны, поиски единства в формах метаболизма, обнаруживающегося в общих биохимических и генетических механизмах, с другой - проникновение в тонкую специфику физиологии отдельных групп микробов. Известные в настоящее время четыре основных типа обмена веществ микроорганизмов - фотолитотрофный, фотоорганотрофный, хемолитотрофный и хемоорганотрофный - детально изучались с помощью различных физико-химических методов.
Хронологически одной из первых проблем, рассмотрение которой дало начальные сведения о физиологических особенностях микробов, было изучение хемоорганотрофного (гетеротрофного) обмена.
После провозглашения Л. Пастером физиологической теории брожения химизм и энзимология процессов брожения стали предметом исследований английской, немецкой, американской и русской школ биохимиков и микробиологов.
Изучение биохимии различных брожений дало представление о диссимилятивной деятельности микроорганизмов и выявило большое разнообразие форм метаболизма в мире микробов.
Решающее значение для выяснения биохимической сущности процессов диссимиляции имела разработка Э. и Г. Бухнерами* (1897) метода бесклеточного спиртового брожения.
* (За эту работу Э. Бухнеру была присуждена в 1907 г. Нобелевская премия.)
Продолжая работы Бухнеров, английские биохимики А. Гарден и В. Юнг (1904) сообщили об успешном разделении бесклеточного дрожжевого сока на две фракции, каждая из которых в отдельности не обладала способностью сбраживать глюкозу. Это открытие повлекло другое, не менее важное: было установлено, что сбраживание углеводов сопровождается образованием фосфорилированных производных, которые Гарден* и Юнг отнесли к промежуточным продуктам процесса брожения. Таким образом была открыта физиологическая функция фосфора в процессах диссимиляции углеводов.
* (За расшифровку механизма брожения углеводов А. Гарден совместно с Г. Эйлером-Хелпином были удостоены в 1929 г. Нобелевской премии.)
В результате этих исследований Гарден и Юнг вывели (1908) уравнение спиртового брожения:
В 1905 г. Л. А. Иванов независимо от Гардена и Юнга пришел к тем же выводам, установив, что прибавление неорганического фосфата к дрожжевому соку ускоряет сбраживание глюкозы, причем фосфат из сока исчезает. В 1911 г. К. Нейберг с помощью разработанного им метода "ловушки" впервые выделил промежуточный продукт распада глюкозы - ацетальдегид. В 1913 г. А. Н. Лебедев разработал эффективный метод получения бесклеточного дрожжевого сока - метод мацерации, обеспечивающий успешное исследование механизма разложения углеводов. Дальнейшему прогрессу в этой области способствовало изучение биохимии и биоэнергетики окисления гликогена в мышцах школой О. Мейергофа (Нобелевская премия, 1922), Г. Эмбденом и Я. О. Парнасом (см. об этом также в главах 3 и 6).
Изучение природы и механизма действия ферментов аэробной и анаэробной диссимиляции углеводов О. Варбургом (Нобелевская премия, 1931) и Г. Эйлером привело в конце 30-х годов к построению схемы, отражающей в основных чертах биохимию гликолиза. Она получила название схемы Эмбдена - Мейергофа - Парнаса. Ее окончательная детализация завершилась в 50-х годах.
Применение ферментных препаратов и радиоактивных изотопов позволило установить наличие у микроорганизмов и других механизмов разложения углеводов. Было выяснено, что реакции, характерные для схемы Эмбдена - Мейергофа - Парнаса, встречаются также и при других бродильных и окислительных процессах.
Одно из первых подтверждений достоверности этих положений дало детальное изучение механизма молочнокислого брожения. Для той формы брожения, которое сопровождается образованием только молочной кислоты, А. Клюйвер и Г. Донкер (1924) предложили термин "гомоферментативное брожение", противопоставив его "гетероферментативному брожению", завершающемуся образованием еще летучих кислот, этилового спирта и СО2. Механизм микробного молочнокислого брожения по аналогии с мышечным гликолизом первоначально рассматривался как гликолиз типа Эмбдена - Мейергофа - Парнаса. Исследованиями М. Гиббса с сотрудниками (1950) было показано, что гомоферментативное брожение протекает по гликолитической схеме, а гетероферментативное - по гексозомонофосфатному пути.
Подробное изучение динамики гомо- и гетероферментативного молочнокислого брожения предприняли В. Н. Шапошников и его ученики. В 50-е годы они пришли к выводу, что при гомоферментативном брожении непосредственным предшественником молочной кислоты является не пировиноградная кислота, а глицериновый альдегид. Изучение с помощью энзимологических и изотопных методов брожений, вызываемых клостридиями и некоторыми бациллами, подтвердили наличие в их механизме элементов классического пути брожения (Ч. Веркман, 1937; О. Осборн, 1937; и др.).
Среди брожений, протекающих по гексозомонофосфатному пути, в частности по схеме Варбурга - Диккенса - Хоррекера, наиболее интенсивно изучались брожения, идущие с превращением пентозофосфата в гексозофосфат и с расщеплением пентоз на двух- и трехуглеродные соединения. Кроме того, изучение брожений, вызываемых Pseudomonas, позволило выявить еще одну разновидность брожений гексозомонофосфатного пути, протекающих по схеме Энтнера - Дудорова. Исследованиями К. Ван-Ниля (80-е годы XIX в.), С. Орла-Иенсена (1898) и особенно Г. Вуда и Ч. Веркмана (40-е годы XX в.) был выявлен химизм брожений, характеризующихся множественностью путей преобразования углеродной цепи и гетеротрофной ассимиляцией СОг. Исследование окислительных брожений обнаружило наличие у микроорганизмов способности к циклическим механизмам окисления.
Изучению различных форм диссимилятивной активности микроорганизмов сопутствовало исследование их энергетического обмена. В результате выявления продуктов неполного окисления субстрата в 30 - 40-х годах была установлена связь между энергетическими и конструктивными процессами (Л. Стикленд, Д. Вудс, В. О. Таусон, А. Клюйвер, К. Ван-Ниль). Важным открытием в этом направлении было установление Ф. Липманом (Нобелевская премия, 1953) функции аденозинтрифосфата (АТФ), выступающего в качестве аккумулятора и переносчика биологической энергии (см. также главу 6).
Важные в теоретическом и практическом отношениях исследования динамики процессов брожения были проведены В. Н. Шапошниковым (40 - 60-е годы), создавшим теорию физиологической фвухфазности брожений. Большой интерес представляет разработка Шапошниковым (1944) классификации процессов, вызываемых хемоорганотрофами, по принципу использования ими субстрата в энергетических и конструктивных целях. Подобные исследования сопровождались накоплением фактических данных о разнообразии ферментных комплексов микроорганизмов.

Владимир Николаевич Шапошников (1884 - 1968)
Изучение фотосинтезирующих бактерий
Важным направлением в физиологии микробов в XX в. было изучение способности некоторых групп микроорганизмов к фотосинтезу - фотолитотрофии и фотоорганотрофии. Общебиологическое значение изучения фотосинтеза состояло в установлении биохимии и кинетики этого важнейшею биологического явления.
Начало обнаружению разнообразия физиологических особенностей фотосинтезирующих микроорганизмов было положено открытием в середине XIX в. зеленых бактерий, а несколько позднее - пурпурных. Г. Молиш (1907) выявил способность пурпурных бактерий расти на органических веществах в темноте и отсутствие выделения ими кислорода. А. Будер (1919) и В. Бевендамм (1924) высказали предположение, что фотосинтезирующие микроорганизмы способны как к фотосинтезу, так и к хемосинтезу. Исследования К. Ван-Ниля показали, что фотосинтезирующие микроорганизмы осуществляют фотосинтез в присутствии окисляемых субстратов - минеральных и органических - и без выделения кислорода. Он же составил уравнение бактериального фотосинтеза:
Исследованиями К. Ван-Ниля (1936), а позднее X. Гаффрона, Дж. Фостера и Д. И. Сапожникова было показано, что специфичность бактериального фотосинтеза определяется именно природой доноров водорода (электрона). Органические соединения могут выполнять функции либо источника водорода (электрона), либо углерода, либо обе эти функции одновременно. Те же функции (иногда в присутствии сульфидов и тиосульфатов) могут нести кислоты цикла Кребса.
В ходе изучения пигментов фотосинтезирующих бактерий, начавшегося с открытия в 1952 г. Г. Шахманом, А. Парди и Р. Стениером хроматофоров, было установлено, что они являются мембранными структурами - ламеллами, которые за уплощенную форму были названы С. Менке (1962) тилакоидами. Среди них были дифференцированы ламеллы. стромы и ламеллы гран, в которых сконцентрированы бактериохлорофиллы. Таким образом, в 50 - 60-е годы стало известно, что фотосинтезирующий аппарат микроорганизмов представляет собой фосфолипопротеиновую структуру и содержит пигменты и переносчики электронов, т. е. дыхательную цепь. Иными словами, система энергетического обмена дополнена у них системой фотосинтезирующих пигментов.
Изучение азотфиксирующих бактерий
Видное место в развитии физиологии микроорганизмов заняли исследования азотфиксирующих микроорганизмов.
В 1901 г. М. Бейеринк и в 1903 г. Дж. Липман выделили три аэробных азотфиксатора - Azotobacter chroococcum, A. agile и A. vinelandii. Позднее азотфиксирующая способность была открыта более чем у 80 видов бактерий, у нескольких видов актиномицетов, спирохет, дрожжей и дрожжеподобных организмов, плесневых и микоризных грибов, а также более чем у 40 видов синезеленых водорослей.
Основную роль в открытии столь широкого распространения способности к азотфиксации и в установлении ее биохимии сыграло применение изотопа азота - 15N. Последовательная смена воззрений на химизм азотфиксации была следующей. Еще до открытия возбудителей азотфиксации А. Готье и Ш. Друэн (1888) предположили, что в процессе связывания азота молекулярный азот сначала окисляется в азотистую, а затем в азотную кислоту. В 1893 г. С. Н. Виноградский высказал мысль о том, что азот восстанавливается выделяющимся водородом с образованием аммиака. Открытие аэробного A. chroococcum было косвенным подтверждением возможности окислительного пути.
Более распространенной, однако, оказалась гипотеза о восстановительном характере процесса. При этом предполагалось либо непосредственное связывание азота с дикарбоновыми кислотами, превращающимися в аминокислоты (М. Герлах, Дж. Фогель, 1902; Дж. Липман, 1903); либо, согласно теории Г. Виланда (1922; Нобелевская премия, 1927), присоединение молекулой азота атома водорода с образованием диимида и гидразина; либо, согласно взглядам С. П. Костычева (1925 - 1931), фиксация молекулярного азота азотобактером происходит внеклеточно путем присоединения водорода к азоту при участии восстановительных ферментов; либо, наконец, по С. Н. Виноградскому (1930), молекулярный азот восстанавливается водородом до аммиака (гидрогенизация). Наряду с этим существовала гипотеза (Д. Блом, 1931), что промежуточным продуктом фиксации является гидроксиламин. Эта идея получила затем развитие в лаборатории А. Виртанена (40-е годы; Нобелевская премия, 1945), который полагал, что гидроксиламин, соединяясь с щавелевой кислотой, образует оксим, превращающийся затем в аспарагиновую кислоту. Эта гипотеза была проверена П. Вильсоном (1954) и не получила подтверждения.
В 1941 г. в исследованиях Р. Бёрриса и Ч. Миллера при изучении фиксации молекулярного азота впервые был использован тяжелый изотоп 15N. Эти ученые экспериментально показали, что первым устойчивым продуктом азотфиксации является аммиак. Этот же факт был установлен и на бесклеточных ферментных системах (Л. Мортенсон и др., 1962; Д. Карнахан и др., 1963; А. А. Имшенецкий и др., 1963; и др.). Однако механизм восстановления N2 до NH3 до сегодняшнего дня остается предметом исследований. В настоящее время известно, что участвующие в азотфиксации ферменты представляют собой систему белковых катализаторов, содержащих в своем составе молибден и железо. В клубеньковых бактериях имеется аналогичный фермент - легоглобищ катализирующий перенос кислорода. Для активного функционирования этих ферментов необходим витамин B12 (В. Л. Кретович, В. А. Яковлев и др.).
Важный научный материал был получен также в результате исследований таких почвенно-микробиологических процессов, как денитрификация, нитрификация, окисление серы, разложение различных органических соединений и многих других процессов, представляющих собой различные формы получения энергии в мире микроорганизмов.
Современный этап в развитии микробиологии
Для развития общей микробиологии в последние десятилетия характерна все более углубляющаяся дифференциация, приводящая к выделению новых самостоятельных направлений. Наряду с продолжающимся развитием экологического направления, связанного с изучением разнообразия мира микробов, принципов и методов их систематики, изучением морфологических, цитологических и цитохимических особенностей микробной клетки шло развитие физиологии микроорганизмов. Оно касалось не только изучения уже названных физиологических свойств микроорганизмов на клеточном, субклеточном и молекулярном уровнях, но и способности отдельных групп микроорганизмов синтезировать биологически активные вещества, белки и аминокислоты, а также подвергать молекулярной трансформации сложные органические соединения.
Следует отметить, что увлечение исследованиями биохимии микробиологических процессов привело к некоторому стиранию граней между биохимией, физиологией и общей микробиологией, к утрате последней ее научной специфики и конкретности в определении научных задач. Оно сопровождалось также некоторым отвлечением внимания ученых от выяснения специфики микроорганизмов как живых биологических объектов. Сложившаяся в общей микробиологии ситуация явилась предметом детального обсуждения на XI Международном микробиологическом конгрессе в Мексике (1970).
Практическое использование биосинтетической и трансформирующей деятельности микробов
С начала второй половины XX в. изучение синтетической и трансформирующей деятельности микроорганизмов ведется в тесном контакте с их селекцией и широким практическим использованием.
Главными продуктами биосинтетической активности микроорганизмов являются белки, витамины, гиббереллины, полисахариды, аминокислоты, ферменты, энтомопатогенные препараты, кормовые антибиотики. Непременным условием успешного развития этого направления стало ведение селекционной работы - получение и использование высокоактивных штаммов продуцентов, обеспечивающих рентабельность производства. Развитие селекции опирается на теоретический фундамент генетики. За сравнительно короткий срок (примерно 20 лет) при помощи селекционно-генетических методов были созданы многие высокоактивные штаммы микробов, продуктивность которых была повышена в 10 - 200 раз по сравнению с исходными штаммами. Их использование явилось предпосылкой создания ряда отраслей микробиологической промышленности. Классическим объектом селекции стали актиномицеты и грибы.
Начало изучения и использования биосинтетической деятельности микроорганизмов связано с получением пенициллина, который в 1940 г. Г. Флори и Э. Чейн выделили из культуры плесневого гриба, впервые описанного А. Флемингом в 1929 г. (совместная Нобелевская премия, 1945). Этот продуцент был идентифицирован М. Тома как Penicillium notatum. Однако родоначальником всех высокоактивных штаммов продуцентов пенициллина, используемых в настоящее время, стал P. chrysogenum.
Существенными нововведениями в производстве пенициллина, значительно повысившими его выход, были: переход с поверхностного выращивания гриба на глубинное (этот новый тип промышленного культивирования приобрел большое значение не только для пенициллиновой, но и для всей микробиологической промышленности); перевод процесса биосинтеза с синтетических питательных сред на богатые питательными веществами среды; введение в ферментационную среду предшественника продукта биосинтеза - осколка пенициллиновой молекулы - фенилуксусной кислоты. Все эти факторы значительно усилили физиологическую активность продуцентов терапевтически наиболее ценного типа пенициллина - бензилпенициллина.
Культура актиномицета - продуцента стрептомицина была впервые выделена в 1914 г. А. Краинским, который подробно описал морфологические и физиологические особенности этого гриба, назвав его Actinomyces griseus. В 1916 г. 3. Ваксман и Р. Куртис выделили другой актиномицет, сходный с предыдущим, отнеся его также к виду Act. griseus. После этого поиски новых продуцентов антибиотиков были возобновлены только в 1943 г., уже после открытия пенициллина и после того, как в 1940 г. 3. Ваксман и X. Вудраф обнаружили у двух видов актиномицетов способность образовывать антибиотические вещества. В 1944 г. Ваксман выделил еще две культуры, продуцирующие стрептомицин (Нобелевская премия, 1952).
В 40 - 50-е годы у микроорганизмов была открыта способность синтезировать и другие антибиотики: окситетрациклин (Act. rimosus, 1948), аурэомицин (биомицин; Str. aureofaciens, 1948), эритромицин (Act. erythreus, 1952), олеандомицин (Act. antibioticus, 1956), грамицидин (Вас. brevis) и т. д. По отношению ко всем продуцентам названных антибиотиков, а также таких антибиотиков, как ванкомицин, ристомицин, канамицин, гризеофульвин, альбомицин, целикомицин, для повышения активности продуцента были применены различные методы селекции.
Столь же успешно исследовалась способность микроорганизмов к синтезу аминокислот. Начало промышленного микробиологического синтеза аминокислот относится к 60-м годам, когда в Японии в результате обработки УФ-лучами исходных штаммов Micrococcus glutamicus, выделенного из почвы (С. Киношита, 1956), были получены штаммы, обладавшие высокой биосинтетической активностью. Изучение механизма синтеза аминокислот, вопросов взаимозаменяемости и конкурентности природных аминокислот и их аналогов позволило значительно глубже проникнуть в содержание физиолого-биохимических процессов, протекающих в микробных клетках. Были установлены явления ретроингибирования, аллостерического торможения при образовании аминокислот, определены места блокирования отдельных этапов биосинтеза у биохимических мутантов, установлены взаимные связи и перекрещивания синтезов различных аминокислот, роль предшественников и т. д.
Изучение биосинтетических путей образования аминокислот микроорганизмами и их селекция интенсивно проводились в течение последнего десятилетия в лабораториях многих стран мира. Микробиологическим путем стали получать аланин, аспарагиновую и глутаминовую кислоты, лизин, метионин, триптофан, лейцин, изолейцин и т. п. По данным С. Киношита (1959) и Н. А. Красильникова (1963), чаще всего в природе встречаются продуценты глутаминовой кислоты и аланина. Анализ путей синтеза аминокислот в клетках показал, что для микроорганизмов характерно восстановительное аминирование пировиноградной кислоты при использовании таких источников углерода, как глюкоза, кислоты цикла Кребса, глицерины, углеводороды; источником аминогруппы является глутаминовая кислота. Образование аланина связано с функционированием аланиндегидрогеназы, обладающей специфичностью по отношению к субстрату и по ответной реакции на воздействие стимулирующих и инактивирующих агентов.
Аспарагиновая кислота, играющая исключительно важную роль в процессах обмена, довольно часто, хотя и в небольших количествах, обнаруживается в культуральной жидкости многих микроорганизмов. Для аспарагиновой кислоты, так же как и для глутаминовой, валина и аланина, характерен синтез путем использования предшественников кетоаналогов и их последующего перевода в аминокислоты ферментными системами клеток. В Японии налажено промышленное производство аспарагиновой кислоты микробиологическим методом на основе превращения фумарата в аспарат Е. coli. При изучении образования глутаминовой кислоты у Pseudomonas ovalis были обнаружены два самостоятельные пути ее синтеза - восстановительное аминирование α-кетоглутаровой кислоты и переаминирование.
Наиболее рациональным способом промышленного биосинтеза лизина оказалось использование мутантов с наследственными нарушениями в цепи превращений аспарагиновой кислоты - исходного продукта для биосинтеза многих аминокислот - лизина, метионина, треонина, изолейцина. В результате сочетания биохимических и генетических методов исследования были получены штаммы, синтезирующие 25 - 30 мг/мл лизина. Среди биохимических методов производства лизина наиболее широкое применение получил способ исключения из числа возможных его предшественников гомосерина и треонина, а также диаминопимелиновой (ДАП) кислоты - основного предшественника лизина, образующегося путем ее декарбоксилирования. Существенным также оказалось использование мутантов с пониженной требовательностью к аэрации.
Большое внимание уделялось изучению путей микробиологического синтеза кормового белка. В качестве субстрата для выращивания продуцентов - главным образом дрожжей рода Candida - используются углеводороды и гидролизаты растительных отходов. Не менее интенсивно велись поиски возможности использования биосинтетической деятельности микробов для получения препаратов различных ферментов. Основными продуцентами ферментов являются грибы рода Aspergillus, а также некоторые бактерии и актиномицеты.
В небольшом количестве получают амилолитические и протеолитиче-ские ферменты и пектиназы. Достаточно хорошо изучены условия биосинтеза таких ферментов, как лактаза, целлюлаза и гемицеллюлаза, фибринолитические ферменты, глюкозооксидаза, нуклеодеполимеразы и др.
Активность ферментов повышают подбором штаммов и условий культивирования, а также методами селекции. Работы в этом направлении широко ведутся в СССР, Японии, США и других странах.
Интенсивное развитие получил также микробиологический синтез органических кислот - лимонной, итаконовой, щавелевой, глюконовой,- наиболее активными продуцентами которых являются аспергиллы; гиббереллинов, основным продуцентом которых служит культура Fusarium moniliforme; витаминов - витамина В2 (рибофлавина), витамина В12 и эргостерина. Выяснилось, что способностью к синтезу витамина В2 обладают дрожжи рода Candida, а также некоторые грибы {Ashbya gossypii, Asp. flavus и Asp. niger) и актиномицеты (Act. olivaceus); продуцентами витамина B12 являются пропионовокислые бактерии, Act. olivaceus, а также микробный комплекс метанового биоценоза, практическое использование которого было разработано в нашей стране (В. Н. Букин, В. Я. Быховский, Е. С. Панцхава и др.). Обработка названных продуцентов мутагенными факторами и последующий отбор дали возможность значительно увеличить синтез ими витамина B12.
За последние годы разностороннему изучению и широкому практическому использованию подверглась трансформирующая активность микроорганизмов. Способность некоторых групп микроорганизмов к тонкой трансформации химических соединений, затрагивающей только один-два атома, нашла применение в тех случаях, когда современная химическая технология еще бессильна. Теоретическую основу развития этого важного раздела физиологии микробов составили идеи В. Л. Омелянского, указывавшего на высокую точность и специфичность деятельности микроорганизмов. "Самые разнообразные реакции окисления и восстановления, гидратации и дегидратации, реакции разложения, полимеризации и атомных группировок,- писал Омелянский,- вызываются микроорганизмами с поразительной легкостью, приводя к глубоким изменениям подвергнутого их воздействию субстрата"*.
* (В. Л. Омелянский. Основы микробиологии. СПб., 1908, стр. 180.)
Микробиологическую трансформацию используют главным образом для получения из стероидного сырья растительного происхождения веществ, обладающих фармацевтическими или гормональными свойствами. Поиски в природе микроорганизмов - трансформаторов стероидной молекулы, способных осуществлять тончайшие реакции (гидроксилирования, дегидрогенизации, дезацетилирования, восстановления и т. п.), ведут по двум направлениям: обнаружение в природе таксономических групп, способных производить ту или иную трансформацию, и выделение методами селекции наиболее активных штаммов. Так были найдены штаммы некоторых грибов, способные осуществлять трансформацию с выходом кортизона и гидрокортизона, актиномицеты и микобактерии, применяющиеся для получения преднизона, преднизолона, диакобала, культуры грибов и микобактерии, селективно дезацетилирующие стероидные соединения.
Исследования трансформирующей активности микроорганизмов основывались на углубленном изучении их физиологии и интенсификации нужных ферментативных процессов. Были найдены также коррелятивные связи между морфологическими свойствами и химической активностью и установлена возможность подбором соответствующих условий (например, окислительно-восстановительных) и усилением нужной ферментативной активности микробов направлять процесс трансформации.
Проблема управляемого культивирования
В последние десятилетия XX в. одной из интенсивно развивающихся проблем микробиологии стала проблема управляемого культивирования. Теоретической основой возникновения и развития этого направления явилось детальное изучение большого разнообразия физиологических потребностей микроорганизмов. Результаты физиологических исследований позволили с помощью соответствующих условий культивирования регулировать ход бродильных и ферментационных процессов, накапливать микробную массу с заданными свойствами, воздействовать на ход микробиологических процессов.
Разработка проблемы управляемого культивирования восходит к исследованиям более раннего периода (20-е годы), направленным на поиск методов изменения хода обмена веществ микроорганизмов с помощью разнообразных факторов внешней среды.
Впервые на принципиальную возможность управлять развитием культуры с помощью условий среды указал Г. Клебс (1905), которому удалось таким образом регулировать развитие грибов и водорослей. Существенное значение имели работы М. Кларка (30-е годы), а в СССР - исследования школы Е. Е. Успенского - С. И. Кузнецова (30 - 50-е годы), развивавшие представление о том, что одним из центральных звеньев превращения веществ являются реакции окисления-восстановления. Полученные ими данные легли в основу представлений об управлении обменом веществ микроорганизмов с помощью изменения различных окислительно-восстановительных условий среды (И. Л. Работнова). Важную роль сыграла также теория двухфазности бродильных процессов, разработанная В. Н. Шапошниковым в 50-е годы.
В 50-е годы XX в. учеными разных стран была разработана теория роста и развития микробов (М. Стефенсон, И. Гунсалус, Н. Д. Иерусалимский, Ж. Моно, В. Шеффер, Р. Финн и др.). Было, в частности, показано, что процесс роста культуры - изменение ее биомассы и численности клеток - сопровождается изменениями структуры клеток, их химического состава и физиологической активности и соответствует физиологическому возрасту культуры. Поэтому регуляция физиологического состояния культуры сводится по существу к регуляции скорости ее роста, осуществляющегося под контролем внутриклеточных механизмов, реагирующих на изменение среды.
В свете этих фактов были разработаны теоретические основы управления ростом микробов, опирающиеся на принцип минимума ("узкого места"), т. е. использование функции лимитирующего вещества (Ж. Моно, А. Новик, Л. Сцилард и др.), а также на регулирование величины популяции и накопления продуктов жизнедеятельности. Полученные данные позволили найти практические методы управляемого культивирования микробов, в том числе метод непрерывного, или проточного, культивирования, теоретическое обоснование которого было дано Ж. Моно в 1950 г.
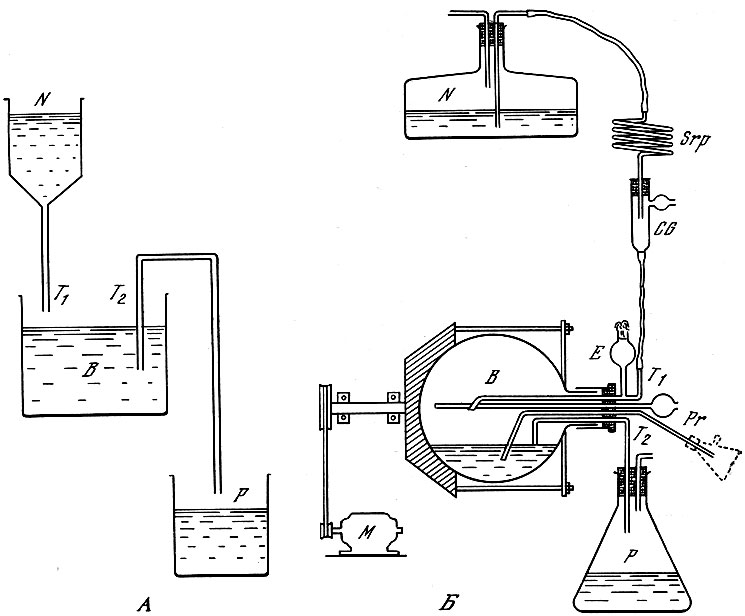
Турбидостат Моно (Б) и схема проточной установки (А) по Моно (1950)
Метод состоит в подаче в культиватор с постоянной скоростью питательной среды и непрерывном выводе с такой же скоростью культуральной среды с бактериями. Точный контроль за непрерывным культивированием микробов осуществляется в аппаратах типа турбидостата, обеспечивающего контроль за плотностью биомассы микроба, либо типа хемостата, позволяющего контролировать скорость увеличения концентрации лимитирующего фактора.
Значительное повышение точности физиологических исследований за счет обеспечения постоянства условий среды и состояния культуры определило широкое использование метода непрерывного культивирования в последние годы.
Основные этапы развития генетики микроорганизмов
В 40 - 50-е годы XX в. из общей микробиологии выделилось самостоятельное направление - генетика микроорганизмов - с наиболее важным разделом - генетикой бактерий. Основой исключительно быстрого развития этой области исследований послужили доказательства мутационной природы изменчивости микроорганизмов, обнаружение у них различных форм генетической рекомбинации и роли ДНК в их наследственных свойствах, достоинства микроорганизмов как объектов генетических исследований. Генетика микроорганизмов приобрела особое значение в связи с решением кардинальных проблем молекулярной биологии. Исследования структуры ДНК, ее роли в процессах биосинтеза белка и регуляции внутриклеточных обменных процессов, разработка проблем направленной изменчивости и специфичности мутагенеза ведутся в основном на микроорганизмах.
Георгий Адамович Надсон (1867 - 1940)
До оформления генетики микроорганизмов в самостоятельную отрасль знания генетики и микробиологи работали разобщенно. Микробиологи были далеки от генетического истолкования наблюдавшихся ими фактов изменчивости и наследственности микробов, а генетики либо вообще не интересовались бактериологией, либо скептически относились к возможности применения генетических принципов к таким примитивно организованным формам жизни, какими им представлялись бактерии.
Первый этап в развитии генетики бактерий составили исследования, экспериментально доказывающие сходство природы и механизмов передачи потомству наследственных признаков у бактерий и высших организмов.
До выделения генетики бактерий в самостоятельную дисциплину существовало несколько точек зрения на популяционную изменчивость бактерий. Согласно одной из них, все изменения, вызванные внешней средой, являются непосредственно адаптивными. Это воззрение основывалось на представлении о наличии у бактерий повышенной врожденной пластичности, позволяющей им адэкватно реагировать на разнообразные условия внешней среды. Длительному сохранению в микробиологии этого взгляда способствовал тот факт, что работа с бактериями велась, как правило, на больших популяциях, быстро размножающихся бесполым путем, что затрудняло контроль за изменчивостью отдельных особей. Вследствие этого изменения наследуемых признаков в единичных клетках оставались не обнаруженными до тех пор, пока такие измененные клетки не размножались в достаточно большом количестве, чтобы образовать целую популяцию.
Согласно другой точке зрения, изменчивость бактерий является результатом длительных модификаций, не затрагивающих наследственной основы организма и проявляющихся в течение многих поколений после того, как вызвавший их фактор перестал действовать. Третья точка зрения сводилась к признанию полицикличности в развитии бактерий, проявлением которой служат изменения их признаков. Эти взгляды означали реставрацию идеи полиморфизма, утверждавшей наличие в мире микробов безграничной изменчивости.
Лишь немногие микробиологи склонялись в четвертой точке зрения, согласно которой причиной изменчивости бактерий могут быть спонтанные изменения (мутации) в одной или нескольких клетках, подвергающиеся отбору. Зачатки подобных представлений содержались, например, в работах М. Нейсера (1906) и Р. Массини (1907), описавших факт внезапного наследственного приобретения культурой Escherichia coli свойства активно сбраживать лактозу. Низкая частота и стабильность этого нового признака позволили авторам получить новый штамм и отнести это явление к категории мутаций, полученных Г. де Фризом. Этим же термином воспользовался и М. Бейеринк при объяснении причин происхождения "дочерних" узелков на поверхности бактериальной колонии. В 1912 г. К. Добелл определил мутации у бактерий как стойкие, иногда незначительные изменения, передающиеся по наследству.
В период с 1910 по 1940 г. были описаны различные виды наследственной изменчивости. В 1921 г. французский микробиолог П. де Крюи описал изменение ряда свойств у одной из патогенных бактерий, возникающее, по его мнению, в результате расщепления признаков под влиянием неблагоприятных условий среды. Этот вид изменчивости он назвал диссоциацией. В 1925 г. в Советском Союзе Г. А. Надсон и Г. С. Филиппов впервые получили мутационные (по их терминологии, сальтационные) стойкие изменения признаков у дрожжевых и плесневых грибков, подвергнув их облучению рентгеновыми лучами. Это был первый случай индуцированной мутации, вызванной физическим мутагенным фактором. Первые данные о характере мутаций позволили провести параллель между природой изменчивости у бактерий и высших организмов. Это стало возможным также после того, как были разработаны методы, которые дали возможность отличать изменения отдельной клетки от изменчивости целой популяции.
Причины различий в объяснении фактов изменчивости у микробов носили преимущественно методологический характер. Из-за отсутствия единой методики эксперимента данные разных авторов оставались несопоставимыми. Трудности в разграничении фенотипа и генотипа приводили зачастую к отрицанию различий между адаптацией и мутацией. Случаи морфологической изменчивости, требующие длительного наблюдения, нередко объясняли наличием сложных циклов развития. Длительное время отсутствовали и единые методы генетического анализа, в частности, принцип отбора мутантов.
Значительное упорядочение представлений о природе изменчивости у микробов было связано с разработкой методов генетического анализа у высших организмов и утверждением представлений о сходстве механизмов изменчивости и наследственности у всех живых организмов.
Генетика бактерий
В генетике бактерий ситуация сложилась таким образом, что, несмотря на достаточное знакомство ученых с явлением спонтанной ненаправленной изменчивости у бактерий, прошло немало времени, прежде чем теория мутаций получила признание. До 40-х годов исследованию подвергалась мутабильность различных признаков бактерий. Преимущественным методом было наблюдение фактов изменчивости и их статистическая обработка. Среди признаков, которые использовались для изучения закономерностей изменчивости, учитывались главным образом антигенность, вирулентность и морфологические особенности. С начала 40-х годов, после признания за мутационной изменчивостью ведущей роли в изменчивости бактерий, характер спонтанных и индуцированных мутаций стали изучать при исследовании таких признаков, как устойчивость к ингибиторам (особенно к антибиотикам), к фагу, потребность в дополнительных факторах роста или дефектность синтеза отдельных ферментов, изменение морфологии клеток или колоний, вирулентные и антигенные свойства. Общему признанию ведущей роли мутационной изменчивости у бактерий способствовала разработка ряда селективных методов ее выявления. Одним из них был метод "флуктуационного теста", разработанный в 1943 г. С. Луриа и М. Дельбрюком*. Метод основывался на учете возникновения фагоустойчивых клонов в популяции чувствительного к фагу штамма Escherichia coli. Его использование положило начало современной генетике бактерий.
* (За работы по генетике бактерий и вирусов С. Луриа и М. Дельбрюку совместно с А. Херши была присуждена в 1969 г. Нобелевская премия.)
Еще более простое доказательство значения мутационной изменчивости дал "метод перераспределения" клеток, разработанный X. Ньюкомбом (1949). Он основан на анализе роли фага при орошении фагофильтратом посевов фагочувствительных бактерий и на учете появления большего по сравнению с контролем числа устойчивых к фагу клеток за счет возникновения мутантных фагоустойчивых клеток.
Веское доказательство существования "преадаптивной" изменчивости было получено методом "отпечатков" бактерий (выросших на чашах-репликах), созданным Дж. Ледербергом* и Э. Ледерберг в 1952 г. Этот метод позволил выявлять спонтанные мутации без обработки фагом или антибиотиками, т. е. в отсутствие специфических условий внешней среды. Еще более наглядно спонтанность возникновения мутантов была продемонстрирована с помощью метода непрямого отбора, разработанного в 1955 г. Л. Кавалли-Сфорца и Дж. Ледербергом. В его основе лежит процесс разведения и обогащения исходной популяции, дающий практически чистую культуру. Данные, полученные с помощью указанных методов, убедили большинство исследователей в том, что основная доля бактериальных мутантов возникает за счет спонтанных и ненаправленных мутаций, частота которых сравнительно низка.
* (За открытие зависимости синтеза ферментов от генетических факторов Дж. Ледерберг совместно с Дж. Бидлом и Э. Тейтумом были удостоены в 1958 г. Нобелевской премии.)
Важным событием в изучении природы мутаций было открытие биохимических (ауксотрофных) мутантов, т. е. микробов, потерявших в результате мутации способность самостоятельно синтезировать те или иные метаболиты и потому нуждающихся во включении этих метаболитов в питательную среду. Ауксотрофные мутанты были впервые обнаружены Дж. Бидлом и Э. Тейтумом в 1941 г. у плесневого грибка Neurosроrа crassa при облучении культуры ультрафиолетовыми лучами. Вслед за тем Дж. Ледерберг и Э. Тейтум (1946) обнаружили у отдельных мутантных клеток Escherichia coli утрату способности синтезировать некоторые аминокислоты и витамины.
Теория Бидла и Тейтума, выраженная формулой "один ген - один фермент", обобщила данные изучения механизмов, контролирующих образование ферментов, с помощью ауксотрофных мутантов. Впоследствии она была уточнена и в настоящее время формулируется как "один ген - одна макромолекула" (РНК или "полипептид"). Важность этих исследований определилась тем, что впервые была установлена связь между отдельным геном и конкретной химической реакцией, происходящей в клетке. Ауксотрофные мутанты стали успешно применяться и при разработке биологических методов определения различных аминокислот, витаминов, азотистых оснований. Как показали исследования Ф. Райяна и Л. Шнейдера в конце 40-х годов, признак потребности в факторах роста может служить маркером как при исследовании динамики популяций, так и при анализе проблемы передачи генетического материала. Исследования с применением ауксотрофов положили начало биохимической генетике.
Восстановление биосинтетической активности у ауксотрофов (или реверсия к прототрофности) было впервые описано Ф. Райяном и Дж. Ледербергом в 1948 г. у Neurosроrа crassa. Это явление рассматривалось как результат либо реверсной мутации в том же локусе, либо мутации в другом локусе хромосомы, сцепленном с локусом, затронутым прямой мутацией (супрессорная мутация). Последующее изучение явления обратного мутирования у биохимических мутантов позволило уточнить механизмы этого процесса. Было показано, что ревертирование обязано не истинным обратным мутациям, а главным образом супрессорным мутациям, возникающим в другом месте генома и приводящим к восстановлению дикого фенотипа. Важный материал был получен также при изучении генетических факторов, регулирующих обмен углеводов, а в связи с этим и механизмов, контролирующих образование ферментов, а также мутирование морфологических признаков клеток, антигенных и вирулентных свойств. Данные по изучению этих мутантов в значительной мере взаимосвязаны (П. де Крюи, 1921; А. Александрини, 1931; В. Браун, 40-е годы).
Для раскрытия молекулярной сущности мутагенеза как главного механизма изменения наследственной информации решающее значение имела расшифровка в 1953 г. структуры молекулы ДНК Дж. Уотсоном и Ф. Криком (Нобелевская премия, 1962) (см. главу 23). Это фундаментальное открытие заложило основу изучения механизмов передачи наследственной информации у бактерий с помощью методов молекулярной биологии. Речь идет об исследовании трансформации, трансдукции, конъюгации и лизогенной конверсии.
Изучение трансформации, трансдукции, конъюгации и лизогенной конверсии
В настоящее время трансформацией называют процесс переноса информации с помощью ДНК от клетки-донора к клетке-реципиенту и замещения в последней в результате рекомбинации специфической последовательности нуклеотидов генома. Термин "трансформация" появился в 1928 - 1934 гг., когда было обнаружено, что некоторые штаммы бактерий, выращенные в присутствии убитых клеток или в культуральных фильтратах и экстрактах из других родственных штаммов, могли приобретать некоторые свойства этих штаммов. Открытие трансформации связано с исследованиями Ф. Гриффита, сообщившего в 1928 г., что при введении мышам смеси живых бескапсульных невирулентных пневмококков (типа II) и убитых нагреванием капсульных вирулентных пневмококков (типа I) они погибали. Из крови мышей были выделены живые капсульные вирулентные пневмококки (тип I), появившиеся в результате превращения невирулентных пневмококков в вирулентные. Это же явление при выращивании клеток R-типа (бескапсульные шероховатые колонии) в бульоне с антисывороткой против этого типа и убитыми нагреванием клетками было воспроизведено in vitro М. Даусоном и Р. Сиа (1931), а вслед за ними Д. Алловеем (1932), но уже не с живыми микробами, а с бесклеточными экстрактами из бактерий, имеющих S-тип (капсульные, слизистые) колонии. Сущность этого явления была выяснена в 1944 г. О. Эвери, К. Мак-Леодом и М. Мак-Карти, установившими, что трансформирующим агентом является высокомолекулярная ДНК (см. главу 23).
Дальнейшее изучение трансформации проводилось при учете в качестве маркеров различных признаков: капсулообразования (для пневмококков), устойчивости к антибиотикам, потребности в факторах роста (X. Раппапорт, 1959; Л. Толмач, Р. Херриотт, 1962; и др.). Исследования с использованием последних двух маркеров дали возможность детально разработать проблемы множественной трансформации и "сцепления" различных генов. Важное место среди этих исследований заняли работы японских ученых X. Иошикавы и Н. Сеока, которые в 1963 г. разработали метод картирования сцепленных генов у Bacillus subtilis.
Изучение динамики процесса трансформации показало, что частота появления трансформантов зависит от физиологического состояния реципиентных клеток. Наибольшая чувствительность (компетентность) клеток к трансформирующему агенту появляется в конце логарифмической и начале стационарной фаз, а затем снижается. Исследованиями М. Фокса и Р. Хочкисса (1957) на пневмококках была выявлена зависимость степени компетентности клеток реципиента к ДНК донора от синтеза белка. Было показано также, что число трансформированных клеток увеличивается до определенного максимума при повышении концентрации введенной ДНК, а также зависит от температуры культивирования клеток, рН среды, аэрации, наличия некоторых катионов. Исследования с меченым фосфором 32Р в молекуле ДНК позволили Л. Лерману и Л. Толмач (1957) сделать вывод о том, что обязательным условием проникновения в клетку ДНК является ее высокая полимерность и сохранение двухтяжной структуры.
Важным моментом в изучении трансформации у бактерий было также выяснение вопроса о том, каким образом происходит включение генетического маркера, несущего определенный признак, в геном бактериальной клетки и когда начинаются его репликация и проявление. Избрав в качестве такого маркера устойчивость к стрептомицину, Г. Эфрусси-Тейлор и Р. Летарже (1955) показали, что начало репликации этого маркера зависит от момента высева трансформируемой культуры на селективную стрептомициновую среду. Извлечением ДНК из реципиентных клеток в различные сроки после включения трансформирующей ДНК эти исследователи устанавливали время включения генетического маркера в геном реципиентной клетки, т. е. определяли время рекомбинации между фрагментами ДНК-донора и ДНК-реципиента. По данным Хочкисса (1956) и Эфрусси-Тейлора (1960), скорость необходимой для этих процессов интеграции экзогенной ДНК зависит от нескольких факторов - величины фрагментов ДНК, природы нуклеотидов, расположенных вблизи от изучаемого маркера, и т. п.
В период интенсивного развития генетики бактерий внимание ученых привлекли вирусы бактерий - бактериофаги. Знакомство с природой бактериофагов выявило огромную перспективность изучения роли этих агентов в передаче генетических признаков для решения общих проблем генетики. В результате уже к 50-м годам генетика бактериофага по темпам развития опередила генетику бактерий (см. главу 25).
Начало изучения роли бактериофага в осуществлении бактериальных рекомбинаций связано с исследованиями Ж. Борде и М. Чуке, открывших в 1921 г. явление лизогении (см. главу 25). Данное ими определение лизогении как наследственной способности бактерий спонтанно продуцировать бактериофаг при отсутствии экзогенного заражения сохранилось и по сегодняшний день. На основе исследований Ф. Бернета, М. Дельбрюка и особенно А. Львова (Нобелевская премия, 1965), проливших свет на истинную природу явления лизогении, стало возможным открытие Дж. Ледербергом и Н. Циндером (1952) нового способа переноса генетической информации с помощью бактериофага, названного ими трансдукцией. Авторы обнаружили это явление при культивировании двух мутантных ауксотрофных штаммов Salmonella tiphimurium в U-oбразной трубке, разделенной ультратонким пористым стеклянным фильтром. После инкубации фаг, содержавшийся в одном из штаммов (22А) - триптофанзависимый, проникал в другой штамм (2А) - гистидинзависимый и переносил наследственное свойство - способность к росту в отсутствие триптофана.
Развивая далее сделанное открытие, Циндер показал, что в основе трансдукции лежит включение фрагментов лизированной фагом бактериальной клетки в фаговую корпускулу. Молекулярная основа этого процесса состоит в передаче чувствительной бактериальной клетке с ДНК фага фрагмента ДНК лизированной бактерии, который интегрируется с участком бактериального генома. Наряду с этим видом трансдукции, названной П. Хартманом (1957) общей неспецифической трансдукцией, была идентифицирована и так называемая ограниченная специфическая трансдукция. Этот тип трансдукции, как показали М. Морзе и Дж. и Е. Ледерберги (1955 - 1956) на фаге лямбда (А,) лизогенного штамма Е. coli K-12, осуществляется при помощи фагов, ДНК которых соединяется лишь с одним определенным участком бактериального генома.
Как выяснили Ф. Кауфман (1953), М. Демерец и П. Хартман (1959), Е. Энглесберг и Л. Барон (1959), в процессе общей трансдукции с помощью фага Salmonella tiphimurium P22 можно трансдуцировать любой ген из многих единичных или сцепленных друг с другом генов, определяющих, например, питательные потребности, серологические и ферментативные признаки, устойчивость к антибиотикам, вирулентность, наличие жгутиков и т. д. При ограниченной трансдукции трансдуктанты обычно нестабильны и могут выщеплять стабильное нелизогенное потомство. Они оказываются, таким образом, не рекомбинантными, а гетерогеномными.
Дальнейшее изучение трансдукции позволило Б. Стокеру (1953), а затем Дж. Ледербергу (1956) описать так называемую абортивную трансдукцию, характеризующуюся тем, что перенесенный признак (фермент) при каждом клеточном делении наследует только одна из дочерних клеток. Как показали исследования с дефектными фагами (В. Арбер, Г. Колленбергер, 1958), при абортивной трансдукции фрагмент бактериальной ДНК, захваченный фагом, вводится в бактерию-реципиент и функционирует в ней, но не интегрируется и не реплицируется с бактериальным геномом. Использование всех трех форм трансдукции способствовало решению многих сложных генетических проблем.
Изучение третьего вида рекомбинаций у бактерий - конъюгации - началось с исследований Дж. Ледерберга и Э. Тейтума, которые в 1946 г. впервые получили положительные результаты при совместном культивировании мутантных ауксотрофных штаммов Е. coli, не способных к росту на минимальной среде. Авторы выделили те клетки, которые в результате рекомбинации получили способность к росту на такой среде и проявляли признаки обоих родительских типов.
В 50-х годах Б. Хейс установил, что при бактериальном скрещивании наблюдается полярность, причем один из партнеров каждой конъюгирующей пары является донором, или "самцом", а другой - реципиентом, или "самкой", т. е. партнеры, участвующие в конъюгации, гетероталличны. Им были выделены два половых типа - F+ (донорные, или мужские штаммы) и F- (реципиентные, или женские штаммы). В 1956 г. Дж. Ледерберг представил прямые микроскопические доказательства образования конъюгационных пар в смешанных культурах рекомбинирующих штаммов.
До 1960 г. было принято считать, что конъюгация по своей природе аналогична половому процессу высших организмов и заключается в слиянии двух гаплоидных клеток с образованием полноценной диплоидной зиготы, которая в результате редукционного деления дает начало гаплоидным рекомбинантным клонам. Позднее в результате исследований Ф. Жакоба, Ж. Моно и их сотрудников - Е. Вольман и Б. Хейс было показано, что, несмотря на гомологичность конъюгации и полового процесса, первая отличается специфическими особенностями, главная из которых состоит в неполной передаче хромосомного материала, благодаря чему образуется частичная диплоидная зигота, или, по терминологии Жакоба и Вольман, мерозигота. Отсюда весь процесс был назван меромиксисом.
В 1963 г. С. Луриа дифференцировал три специальных генетических фактора, необходимых для установления клеточных контактов при конъюгации, которые он назвал "конъюгонами": F - факторы фертильности, сf - факторы колициногенности - генетические структуры, которые передаются при клеточном контакте и являются ответственными за образование колицинов - антибиотиков полипептидной природы, образуемых некоторыми штаммами Е. coli, и RTF - фактор передачи устойчивости к антибиотикам. При наличии в клетке-доноре любого из этих факторов может происходить конъюгация с родственной клеткой, не несущей такого фактора.
Дальнейшие исследования на молекулярном уровне показали, что по своей химической природе конъюгоны являются ДНКсодержащими элементами; они имеют, вероятно, кольцевую структуру, способны к самоудвоению и связаны с клеточной мембраной.
Приведенные представления о природе конъюгонов находятся в тесной связи с открытием в 1958 г. Жакобом и Вольман эписом - дополнительных внехромосомных генетических элементов, которые могут находиться как в автономном, так и в интегрированном с хромосомой состоянии. Ввиду накопления данных, свидетельствующих о том, что генетические элементы типа F и сf-конъюгонов отличаются от ДНКсодер-жащих структур ("хромосом") лишь по величине и темпам репликации, Ф. Жакоб и С. Бреннер (1963) предложили для них общее название - репликоны. Как теперь известно, нехромосомные репликоны, или эписомы, служат передатчиками генетической информации при вирусной конверсии, сущностью которой является привнесение с помощью фага дополнительной информации, содержащейся в фаговой ДНК, которая в рекомбинации не участвует; изменения же некоторых наследственных признаков происходят за счет этих функционирующих битов информации. При этом изменения наблюдаются лишь до тех пор, пока фаговая ДНК, внесенная внутрь данной клетки, сосуществует с бактериальной.
Явление генетической рекомбинации у актиномицетов впервые описал в 1955 г. Г. Сермонти. Двумя годами позднее С. И. Алиханян и С. 3. Миндлин сообщили об использовании биохимических мутантов Actinomyces rimosus для получения гибридных форм. Однако механизм осуществления рекомбинаций у актиномицетов оставался до самого последнего времени неясным. В настоящее время можно считать установленным, что механизм генетической рекомбинации у актиномицетов принципиально сходен с таковым у бактерий. В то же время актиномицеты с их сложной морфологической организацией стали ценным объектом генетики микроорганизмов. В результате генетических исследований было показано, что актиномицеты хорошо растут на простых синтетических средах и это обеспечивает получение у них разнообразных ауксотрофных линий; хорошо спорулируют, образуя в большинстве случаев легко отделяющиеся друг от друга гаплоидные споры; подобно бактериям, у них имеются специфические фаги (актинофаги) и известно явление лизогении. Актиномицеты служат продуцентами подавляющего большинства антибиотиков и классическими объектами в селекции промышленных микроорганизмов.
Методы генетического анализа рекомбинантов у актиномицетов были разработаны Дж. Хопвудом и Г. Сермонти (1962). Один из них основан на анализе гаплоидных рекомбинантов, другой - на анализе в гетероклонах (гетерогенные колонии, которые вырастают не из гаплоидных спор, как обычные гаплоидные рекомбинанты, а из диплоидных или частично диплоидных, в связи с чем они не прорастают на селективных для гаплоидных рекомбинантов средах). Образование гетероклонов, т. е. потомства мерозигот, несущих помимо генома одного из родителей часть генома другого родителя, свидетельствует о сходстве генетической рекомбинации бактерий и актиномицетов. Принципиальной особенностью анализа в гетероклонах оказалась возможность определения величины сцепления между генетическими локусами в абсолютных единицах. Согласно гипотезе, выдвинутой этими учеными, для гетероклонов характерна неполная диплоидность ядра: они происходят из неполных гетерозигот, в которых отсутствует один или два конечных сегмента хромосомы в каждой группе сцепления. Исследования Хопвуда на Str. coelicolor (1965) послужили доводом в пользу представления не о линейных группах сцепления локусов, а о единой кольцевой группе, в результате чего жизнеспособные рекомбинанты возникают только благодаря четному числу перекрестов. Эта же особенность сцепления свидетельствует о сходстве организации генетического материала у бактерий и актиномицетов. В то же время отсутствие четких результатов по передаче и наследованию фактора фертильности (фактора F), а также отсутствие штаммов Hfr не позволяет еще полностью гомологизировать системы половой полярности у бактерий и актиномицетов.
Впервые о возможности генетической трансдукции у актиномицетов сообщили в 1959 г. С. И. Алиханян и Т. С. Ильина, которые в 1960 г. описали трансдукцию с помощью актинофага, выделенного из штамма Act. olivaceus, нуждающегося в метионине, гистидине и цистине. В отличие от бактерий, трансдуктанты у актиномицетов имеют не один, а два типа. В свою очередь рекомбинация у актинофагов, проявляющаяся в основном в изменении их литических свойств, так же как и у бактериофагов, была описана в 1969 г. С. И. Алиханяном и Н. Д. Ломовской.
Начало изучению рекомбинаций у несовершенных грибов было положено открытием в начале 50-х годов Г. Понтекорво у Aspergillus nidulans явления, названного парасексуальным процессом. Механизм этого процесса, как было показано, не отличается по существу от механизма рекомбинации при настоящем половом процессе. X. Хансен (30-е годы), Г. Понтекорво (1946), Дж. Л. Джинкс (1952), Дж. А. Ропер (1952) и другие установили, что парасексуальный цикл у грибов состоит из трех последовательных этапов: образования гетерокарионов - форм, совмещающих ядра двух разных типов; возникновения в мицелии гетерокариона гетерозиготных диплоидных ядер в результате слияния двух ядер разных типов; образования рекомбинантных форм благодаря митотическому расщеплению. В настоящее время известно, что парасексуальный процесс довольно широко распространен среди грибов. Механизм его может отличаться у различных видов; он может быть использован как при генетическом анализе, так и при селекции видов, представляющих практический интерес.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'