2.2.4. Рост и размножение
Индивидуальный рост и бесполое размножение клеток
Отношение поверхность/объем у бактериальных клеток очень велико. Это способствует быстрому поглощению питательных веществ из окружающей среды за счет диффузии и активного транспорта. В благоприятных условиях бактерии растут очень быстро. Рост прежде всего зависит от температуры и рН среды, доступности питательных веществ и концентрации ионов. Облигатным аэробам обязательно нужен еще и кислород, а облигатным анаэробам, наоборот, нужно, чтобы его совсем не было. Достигнув определенных размеров, бактерии переходят к бесполому размножению (бинарному делению), т. е. начинают делиться с образованием двух дочерних клеток. Переход к делению диктуется отношением объема ядра к объему цитоплазмы. Перед клеточным делением происходит репликация ДНК, во время которой мезосомы удерживают геном в определенном положении (рис. 2.3 и 2.4). Мезосомы могут прикрепляться и к новым перегородкам между дочерними клетками и каким-то образом участвовать в синтезе веществ клеточной стенки. У самых быстрорастущих бактерий деление происходит через каждые 20 мин; интервал между делениями называется временем генерации.
Рост популяции
2.1. Рассмотрим ситуацию, когда одиночная бактериальная клетка помещена в питательную среду и находится в условиях, оптимальных для роста. Перепишите табл. 2.2. Заполните ее для случая, когда эта клетка и все ее потомки делятся, допустим, каждые 20 мин.
На основе полученных вами данных постройте две кривые. По горизонтальной оси отложите время, а по вертикальной - либо число клеток (кривая А), либо десятичный логарифм этого числа (кривая Б). Что вы можете сказать о форме этих кривых?
Когда число клеток увеличивается, как показано в табл. 2.2, говорят о логарифмическом, экспоненциальном или геометрическом росте. В этом случае мы получаем экспоненциальный ряд чисел. Это гораздо проще понять, если посмотреть на строку В в табл. 2.2, где число бактерий выражено в виде числа 2, возведенного в соответствующую степень. Показатель степени можно назвать логарифмом или экспонентой числа 2. Логарифмы или экспоненты образуют линейный ряд (0, 1, 2, 3 и т. д.), соответствующий числу генераций.

Таблица 2.2. Рост бактерий в модельной популяции
Вернемся к табл. 2.2 и посмотрим, как числа, расположенные в строке А, превращаются в логарифмы по основанию 2:
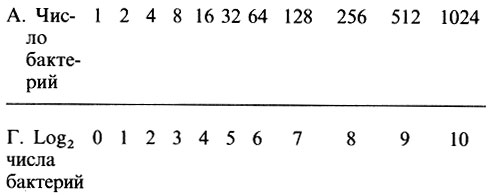
Таблица 2.2. Рост бактерий в модельной популяции
Сравните строки А и Г. Однако обычно пользуются десятичными логарифмами (см. строку Б). В этом случае 1=100, 2=100'3, 4=100'6 и т. д.
Кривая, изображенная на графике А, называется логарифмической или экспоненциальной кривой. Такую кривую можно преобразовать в прямую, построив график изменения логарифма числа клеток во времени. Тогда в идеальных условиях рост бактерий теоретически должен быть экспоненциальным. Сравним эту математическую модель с кривой роста реальной популяции бактерий, которая изображена на рис. 2.7. Можно отметить четыре фазы роста. Во время лаг-фазы бактерии адаптируются к новой среде обитания, и поэтому максимальная скорость роста не достигается. В этот период у бактерий могут, например, синтезироваться новые ферменты, необходимые для усвоения тех питательных веществ, которые содержатся в новой среде.
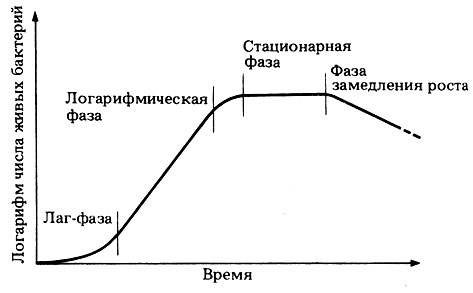
Рис. 2.7. Типичная кривая роста популяции бактерий
Логарифмическая фаза - это такая фаза, когда бактерии растут с максимальной скоростью, число клеток увеличивается почти экспоненциально, а кривая роста идет прямолинейно. Затем рост колонии начинает замедляться, и культура входит в стационарную фазу, когда скорость роста равна нулю и когда резко возрастает конкуренция за пищевые ресурсы. Образование новых клеток замедляется, а затем совсем прекращается. Увеличение числа клеток компенсируется одновременной гибелью других клеток, поэтому суммарная численность живых клеток остается постоянной. Переход к этой фазе обусловлен действием многих факторов: истощением среды, накоплением токсичных "шлаков", образующихся в процессе обмена веществ, а в случае аэробных бактерий еще и уменьшением содержания кислорода в среде.
Во время последней фазы - фазы замедления роста - ускоряется гибель клеток и прекращается их размножение. Способы подсчета числа бактерий описаны в конце этой главы.
2.2. Какую кривую роста мы получим, если возьмем пробу бактерий из культуры, достигшей стационарной фазы роста, перенесем ее в свежую среду и затем оценим рост бактериальной популяции?
2.3. Культура бактерий была помещена в питательный раствор и инкубировалась в нем при 30°С. Сразу же после посева и через интервалы, указанные в табл. 2.3, было определено число бактерий в культуре.
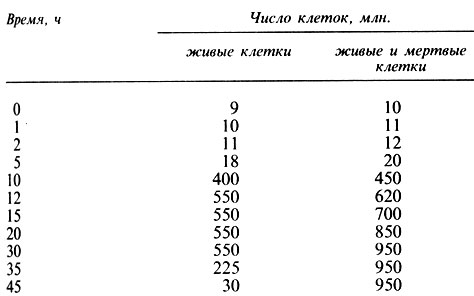
Таблица 2.3. Рост культуры бактерий при 30°С
Используя эти цифры, постройте график и разберитесь, что произошло. Посмотрите на полученные кривые и скажите, чем, по вашему мнению, вызваны наблюдаемые изменения численности бактерий. (Экзаменационная работа "А"-уровня, вопрос 11, Оксфорд, лето 1976 г.)
2.4. Каково "время генерации" бактерий в задаче 2.3?
Половое размножение, или генетическая рекомбинация
У бактерий наблюдается и половое размножение, но в самой примитивной форме. Половое размножение бактерий отличается от полового размножения эукариот тем, что у бактерий не образуются гаметы и не происходит слияния клеток. Однако главнейшее событие полового размножения, а именно обмен генетическим материалом, происходит и в этом случае. Этот процесс называется генетической рекомбинацией. Часть ДНК (очень редко вся ДНК) клетки-донора переносится в клетку-реципиент, ДНК которой генетически отличается от ДНК донора. При этом перенесенная ДНК замещает часть ДНК реципиента. В процессе замещения ДНК участвуют ферменты, расщепляющие и вновь соединяющие цепи ДНК. При этом образуется ДНК, которая содержит гены обеих родительских клеток. Такую ДНК называют рекомбинантной. У потомства, или рекомбинантов, наблюдается заметное разнообразие признаков, вызванное смешением генов. Такое разнообразие признаков очень важно для эволюции и является главным преимуществом полового размножения.
Известны три способа получения рекомбинантов. Это - в порядке их открытия - трансформация, конъюгация и трансдукция.
При трансформации клетки донора и реципиента не контактируют друг с другом. Этот процесс открыл в 1928 г. Гриффит (Griffith), работая с пневмококками-бактериями, вызывающими пневмонию. У пневмококков имеются колонии двух типов, которые различаются по внешнему виду. Одни колонии - шероховатые (R-от англ. rough - шероховатый), другие - гладкие (S-от англ. smooth - гладкий, ровный). R-штаммы не патогенны и не образуют капсулы; S-штаммы патогенны, и у них имеются толстые капсулы (разд. 2.2.2). Гриффит обнаружил, что если мышам ввести живые R-клетки и мертвые (убитые нагреванием) S-клетки, то мыши погибают через несколько дней, а в крови у них можно обнаружить живые S-клетки. На этом основании Гриффит сделал вывод, что из мертвых S-клеток высвобождается какой-то фактор, который придает R-клеткам способность образовывать капсулу и предохраняет их от разрушения в организме животного-хозяина. Оказалось, что такая "трансформация" наследуется. Поскольку молекулы "наследственности" в то время еще не были известны (хотя, правда, и предполагали, что это белки), очень много усилий было потрачено на то, чтобы идентифицировать трансформирующий фактор.
В 1944 г. Эвери, Мак-Леоду и Мак-Карти (Avery, MacLeod, McCarty) удалось выделить и идентифицировать этот фактор. К изумлению исследователей им оказалась ДНК, а не белок. Так были получены первые прямые данные о том, что генетическим материалом является ДНК.
Ныне известно, что при трансформации из клетки-донора выходит небольшой фрагмент ДНК, который активно поглощается клеткой-реципиентом и включается в состав ее ДНК, замещая в ней похожий, хотя и не обязательно идентичный фрагмент. Трансформация наблюдается лишь у немногих бактерий, в том числе и у некоторых так называемых "компетентных" штаммов пневмококков, у которых ДНК может проникать в клетку-реципиент. Возможный механизм трансформации изображен на рис. 2.8.
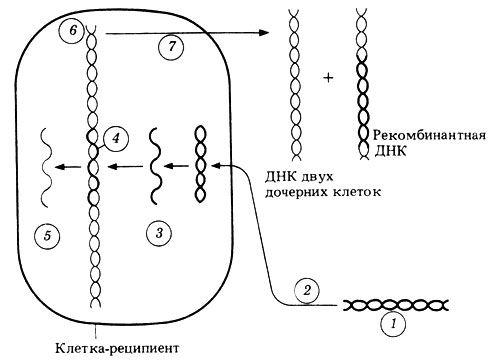
Рис. 2.8. Один из возможных способов трансформации. Точный механизм активного поглощения ДНК донора неизвестен. 1 - ДНК донора; 2 - активное поглощение; 3 - ДНК донора становится одноцепочечной (вторая цепь разрушается); 4 - цепь ДНК донора замещает сходную, но не идентичную цепь ДНК реципиента; 5 - вытесненный фрагмент реципиентной ДНК затем разрушается; 6 - гибридная ДНК; 7 - репликация гибридной ДНК
Конъюгация - это перенос ДНК между клетками, непосредственно контактирующими друг с другом. В отличие от трансформации и трансдукции при этом может обмениваться значительная часть до-норной ДНК. Этот процесс был открыт в 1946 г. у Escherichia coli. Был проведен такой опыт. Обычно клетки Е. coli синтезируют все необходимые им аминокислоты, если в среде содержится достаточно глюкозы и неорганических солей. В результате облучения бактерий иногда образуются мутанты. Были выбраны два мутанта: мутант, не способный синтезировать витамин биотин и аминокислоту метионин, и мутант, не способный синтезировать аминокислоты треонин и лейцин. В среду, не содержавшую все эти четыре фактора роста, помещали по 108 клеток каждого штамма. Теоретически клетки не должны были расти в такой среде. Однако все же было получено несколько сотен колоний (каждая колония возникает всего из одной начальной клетки), причем оказалось, что в таких клетках имеются все гены, необходимые для образования этих четырех факторов роста. Следовательно, произошел какой-то обмен генетической информацией, но выделить вещество, ответственное за этот процесс, в то время не удалось. В конце концов было установлено (при помощи электронного микроскопа), что клетки Е, coli могут непосредственно контактировать друг с другом, т. е. конъюгировать (рис. 2.9).
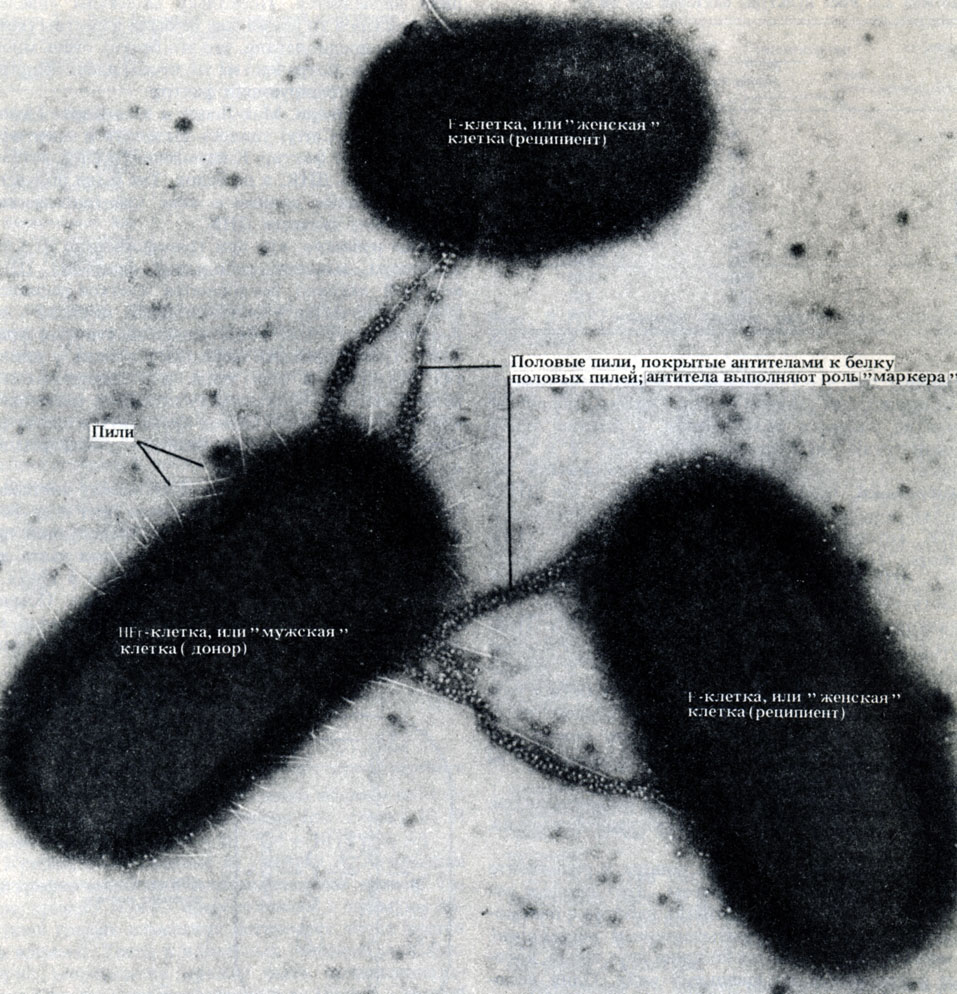
Рис. 2.9. Микрофотография конъюгирующих бактерий (одной 'мужской' и двух 'женских' особей), полученная с помощью просвечивающего электронного микроскопа. × 19475
Донорная способность клеток определяется генами, находящимися в небольшой кольцевой молекуле ДНК, которую называют половым фактором или F-фактором (F - первая буква от англ. fertility - плодовитость). Это - своеобразная плазмида (см. ниже), которая кодирует белок специфических фимбрий, называемых F-пилями или половыми пилями. F-пили облегчают контакт клеток друг с другом. Молекула ДНК состоит из двух цепей. При конъюгации одна из цепей двухцепочечной ДНК F-фактора проникает через половую фимбрию из клетки-донора (F+) в клетку-реципиент (F-). Этот процесс схематически показан на рис. 2.10. Видно, что в клетке-доноре сохраняется F-фактор, который реплицируется в ней, пока в клетке-реципиенте синтезируется ее собственная копия. Так постепенно вся популяция клеток становится F+-клетками. Клетки-доноры могут спонтанно утрачивать F-фактор и становиться, таким образом, F--клетками.
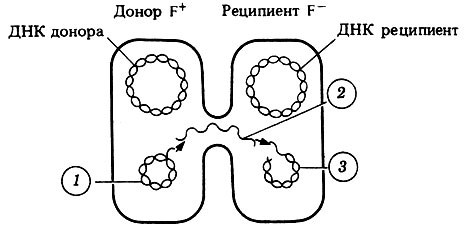
Рис. 2.10. Конъюгация и перенос F-фактора из клетки в клетку. 1, 2 и 3 обозначают последовательность этапов переноса. 1 - раскручивающийся и одновременно реплицирующийся F-фактор; 2 - одноцепочечный F-фактор проникает в клетку-реципиент через F-фимбрию; 3 - F-фактор с синтезирующейся комплементарной цепью
F-фактор интересен еще и потому, что иногда (примерно в 1 случае из 100000) он встраивается в молекулу основной ДНК клетки-хозяина. Тогда при конъюгации переносится не только F-фактор, но также и остальная ДНК. Этот процесс занимает примерно 90 мин, но клетки могут расходиться и раньше, до полного обмена ДНК. Такие штаммы постоянно передают всю или большую часть своей ДНК другим клеткам. Эти штаммы называют Hfr-штаммами (от англ. Н = High - высокая, f = frequency - частота, г т recombination - рекомбинация), потому что донорная ДНК таких штаммов рекомбинирует с ДНК реципиента.
При трансдукции небольшой двухцепочечный фрагмент ДНК попадает из клетки-донора в клетку-реципиент вместе с бактериофагом (одна из групп вирусов, см. разд. 2.5). Возможный механизм трансдукции изображен на рис. 2.11.
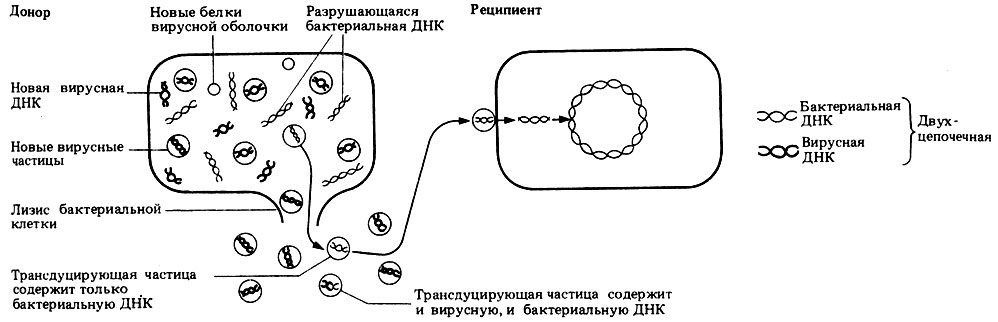
Рис. 2.11. Механизм трансдукции
Некоторые вирусы способны встраивать свою ДНК в ДНК бактерий; такая встроенная ДНК реплицируется одновременно с ДНК хозяина и передается от одного поколения бактерий к другому. Время от времени такая ДНК активируется и начинает кодировать образование новых вирусов. ДНК хозяина (бактерии) разрывается, а высвобожденные фрагменты иногда захватываются внутрь новых вирусных частиц, порой даже вытесняя ДНК самого вируса. Такие новые "вирусы", или трансдуцирующие частицы, затем переносят ДНК в клетки других бактерий.
Плазмиды и эписомы
Плазмиды и эписомы - это небольшие фрагменты ДНК, отличающейся от основной массы ДНК. Они часто реплицируются вместе с ДНК хозяина, но не нужны для выживания его клетки.
Сначала было принято различать эписомы и плазмиды: эписомы внедряются в ДНК хозяина, а плазмиды - нет. К эписомам относятся F-факторы и так называемые умеренные фаги (разд. 2.5.4). Сейчас обе группы называют одним общим термином "плазмиды". Плазмиды широко распространены в природе, и в последние годы их считают внутриклеточными паразитами или симбионтами, устроенными еще проще, чем вирусы. Вопрос о том, можно ли вирусы считать живыми организмами, мы обсудим в разд. 2.5.2. Что касается плазмид, то здесь дело обстоит еще сложнее - ведь они представляют собой только молекулы ДНК.
Плазмиды придают своим клеткам-хозяевам целый ряд особых свойств. Некоторые плазмиды являются "факторами резистентности" (R-плазмиды, или R-факторы)*, т. е. факторами, придающими устойчивость к антибиотикам. Примером может служить пенициллиназная плазмида стафилококков, которая трансдуцируется различными бактериофагами. В этой плазмиде содержится ген, кодирующий фермент пенициллиназу, которая разрушает пенициллин и, таким образом, придает устойчивость к пенициллину. Передача и распространение таких факторов среди бактерий (в результате полового размножения) очень мешают врачам. Другие плазмидные гены определяют устойчивость к дезинфицирующим средствам; способствуют таким заболеваниям, как стафилококковая импетиго; помогают молочнокислым бактериям превращать молоко в сыр; придают способность усваивать такие сложные вещества, как углеводороды, что можно использовать для борьбы с загрязнениями океана или для получения кормового белка из нефти.
* (От англ. R = resistance - устойчивость. - Прим. перев.)
В заключение следует сказать, что половое размножение (в любой форме) - довольно редкое событие у бактерий. Но поскольку число бактерий в каждой колонии огромно, половое размножение наблюдается сравнительно часто. Такое размножение более примитивно, чем у эукариот; полный обмен геномами (суммарной ДНК) происходит только при конъюгации, что действительно встречается лишь изредка. Половое размножение бактерий имеет особое значение потому, что именно таким путем передается устойчивость к антибиотикам и дезинфицирующим средствам.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'