3.2. Водоросли (Algae)
3.2.1. Основные признаки и систематика водорослей
Водоросли - огромная группа растений, имеющих большое биологическое значение и очень важных для человечества (разд. 3.2.8). Они являются самыми примитивными из растений, и у них нет разделения тела на стебель, корень и листья. Поэтому первоначально их объединяли вместе с грибами в отдел Thallophyta (см. примечание на с. 43). Однако после новых научных открытий стало ясно, что водоросли ничуть не менее разнообразны, чем все остальные группы растений, вместе взятые, и что у них очень мало общих признаков. Вероятно, лучше всего считать водорослями все фотосинтезирующие организмы, выделяющие кислород, которые эволюционировали в водной среде и полностью освоили ее. Правда, некоторые водоросли вышли и на сушу, но в масштабах планеты продуктивность прибрежных и наземных форм ничтожна в сравнении с продуктивностью океанических и пресноводных водорослей. Если придерживаться такой точки зрения, то в группу водорослей следует включить и сине-зеленые водоросли (Cyanophyta). Однако, поскольку эти водоросли - прокариоты, было предложено называть их цианобактериями (Cyanobacteria), чтобы как-то отличить от эукариотических водорослей. При этом из виду упускают один очень важный факт, а именно то, что сине-зеленые водоросли при фотосинтезе выделяют кислород, а все остальные фотосинтезирующие прокариоты - нет. Для того чтобы вода могла расщепиться на водород и кислород, необходимо наличие хлорофилла и фотосистемы II (разд. 9.4.2), что является важным преимуществом перед фотосинтезирующими бактериями. О том, как было достигнуто такое преимущество, известно очень мало, хотя и обнаружены некоторые формы, занимающие промежуточное положение между сине-зелеными водорослями и бактериями. Такое истолкование связи между сине-зелеными водорослями и другими растениями, в том числе и остальными водорослями, подкрепляют данные, свидетельствующие в пользу симбиотической теории, согласно которой хлоропласты растений произошли из сине-зеленых водорослей (разд. 9.3.1).
Подводя итог, можно сказать, что термин "водоросль" сам по себе удобен, но его применение в систематике вносит ненужные осложнения. Сине-зеленые водоросли следует относить к прокариотам, а все остальные водоросли - к эукариотам.
К счастью, эукариотические водоросли вполне естественно распадаются на хорошо различимые группы, причем основным отличительным признаком является набор фотосинтетических пигментов. В современной систематике такие группы получили статус отделов. Родственные связи между отделами до сих пор не выяснены, а этот вопрос очень важен, чтобы понять происхождение высших растений и связь между прокариотами и эукариотами.
Все отделы перечислены на рис. 3.11, а на рис. 3.12 даны современные представления о том, какие связи существуют между этими отделами. Основные признаки водорослей и некоторых главных отделов приведены в табл. 3.4.

Рис. 3.11. Систематика эукариотических водорослей
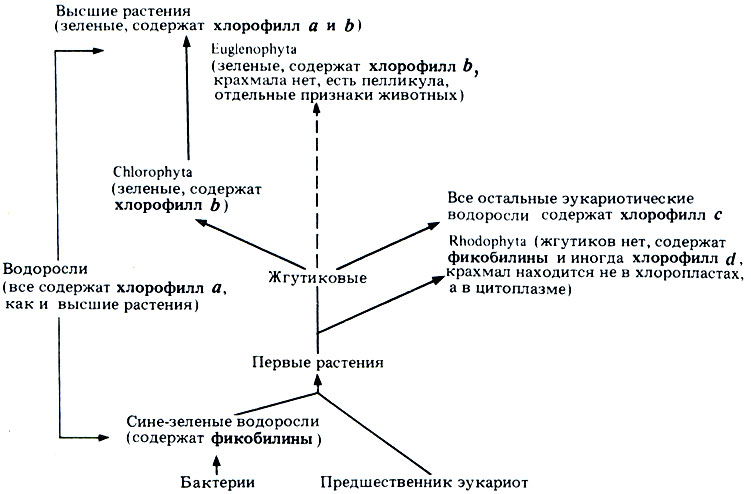
Рис. 3.12. Возможная взаимосвязь между различными группами водорослей, бактериями и высшими растениями. Более подробно этот вопрос рассматривается в работе: В. S. Rushton, School Science Review, 62, № 221, 648-654, June 1981; этот автор выделяет три подцарства, соответствующие наличию хлорофилла d (вместе с фикобилинами), хлорофилла с или хлорофилла b. К последнему подцарству должны тогда относиться Chlorophyta и все высшие растения
3.2.2. Бесполое размножение водорослей
У водорослей наблюдается как бесполое, так и половое размножение. Ниже кратко перечислены основные типы бесполого размножения от самого простого до самого сложного.
Вегетативное размножение. У некоторых колониальных форм колонии могут дробиться на отдельные фрагменты, которые дают начало новым более мелким колониям. У более крупных водорослей, например у Fucus, на главном талломе могут образовываться дополнительные талломы, которые отламываются и образуют новые организмы.
Фрагментация. Это явление наблюдается у нитчатых водорослей, таких, как сине-зеленые водоросли и Spirogyra. Нить расщепляется строго определенным образом вдоль, и образуются две новые нити. Это явление можно рассматривать как одну из форм вегетативного размножения.
Бинарное деление. В этом случае одноклеточный организм делится на две одинаковые половины, при этом ядро делится митотически. Продольное деление такого типа наблюдается у Euglena.
Зооспоры. Это подвижные споры, имеющие жгутики. Они образуются у многих водорослей, например у Chlamydomonas, и у некоторых грибов (см. Oomycota, табл. 3.2).
Апланоспоры. Эти неподвижные споры образуются, например, у некоторых бурых водорослей.
3.2.3. Половое размножениеводорослей
При половом размножении объединяется генетический материал двух отдельных особей одного и того же вида. Самый простой способ такого размножения у водорослей; он заключается в слиянии двух морфологически (т. е. структурно) идентичных гамет. Такой процесс называется изогамией, а гаметы - изогаметами. Изогамны Spirogyra и некоторые виды Chlamydomonas.
Если одна из гамет менее подвижна или крупней, чем другая, то такой процесс называют анизогамией. У Spirogyra гаметы не отличаются по строению, но одна из них двигается, а другая неподвижна. Это можно рассматривать как физиологическую анизогамию. Существует еще один вариант, когда одна гамета большая и неподвижная, а вторая небольшая и подвижная. Такие гаметы называются женскими и мужскими, а сам процесс называется оогамией. Оогамны Fucus и некоторые виды Chlamydomonas. Женские гаметы крупные потому, что в них находится запас питательных веществ, необходимых для развития зиготы после оплодотворения.
Все три типа полового размножения соответствуют увеличению сложности строения тела, и поэтому оогамия, хотя и встречается у некоторых простых водорослей, таких, как Chlamydomonas, в целом более распространена у более сложных водорослей, например у представителей Phaeophyta. Оогамия - это единственный способ полового размножения у растений, более высоко организованных, чем водоросли.
К сожалению, терминология, применяемая для описания гамет и органов полового размножения растений, очень запутана, особенно у водорослей. Ниже мы объясним только основные термины.
У грибов и низших растений (водорослей, мохообразных и папоротникообразных) гаметы образуются в особых структурах, которые называют гаметангиями. Мужской гаметангий называется антеридием, а женский - оогонием или архегонием.
Оогоний* - это простой женский гаметангий, который встречается у многих водорослей и грибов, а женские гаметы или гаметы, которые находятся в нем, называют оосферами. Оплодотворенная оосфера называется ооспорой; она превращается в толстостенную покоящуюся спору, способную переживать неблагоприятные условия. Общее название для женской гаметы - яйцо или яйцеклетка, хотя иногда для обозначения яйцеклетки используют термин "оосфера"; однако это не совсем точно.
* (Оогониями называют также клетки яичников, из которых образуются ооциты у животных (см. гл. 20).)
Архегоний - это более сложный женский гаметангий, который характерен для мохообразных, папоротникообразных и многих голосеменных; архегоний будет описан далее в этой главе.
В антеридии образуются мужские гаметы, которые называются антерозоидами или сперматозоидами. Они подвижны, потому что снабжены одним или несколькими жгутиками. Такие гаметы характерны для грибов, водорослей, мохообразных, папоротникообразных и некоторых голосеменных. У животных мужские гаметы называются сперматозоидами или спермиями. Перечисленные названия приведены на рис. 3.13.
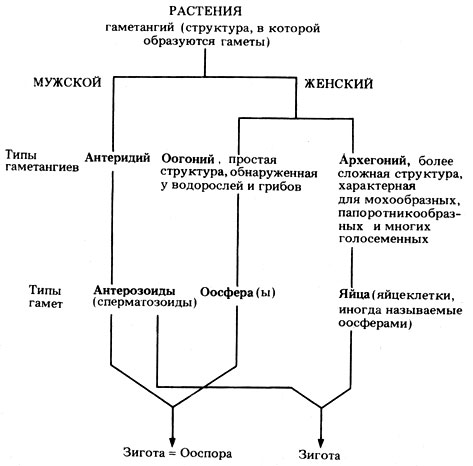
Рис. 3.13. Типы гаметангиев и гамет, встречающиеся в растительном царстве
Для тех целей, которые стоят перед нами в этой главе, не столь важно, как называть разные гаметы одного и того же пола, поэтому вполне достаточно различать сперматозоиды, т. е. все мужские гаметы, и яйцеклетки, т. е. все женские гаметы.
Как и у грибов, у некоторых водорослей наблюдается гетероталличность (разд. 3.1.3).
3.2.4. Отдел Chlorophyta
Основные свойства Chlorophyta перечислены в табл. 3.4.

Таблица 3.4. Систематика и основные признаки некоторых главных групп водорослей1)
1) (Звездочкой отмечен систематический признак.)
Chlamydomonas (хламидомонада) - одноклеточная подвижная водоросль, которая живет главным образом в стоячей воде, т. е. в прудах и канавах, особенно если вода еще и обогащена растворимыми азотистыми соединениями, например стоками со скотных дворов. Клетки этой водоросли часто встречаются в таком огромном количестве, что вода становится зеленой. Некоторые виды живут в морской воде или в солоноватых лиманах.
Строение
Хламидомонада совсем не похожа на растение, так как она активно движется и у нее имеются пульсирующие вакуоли. Строение хламидомонады изображено на рис. 3.14. На электронной микрофотографии видны типичные для эукариот органеллы: аппарат Гольджи, митохондрии, рибосомы и мелкие вакуоли. В хлоропластах многих водорослей выявлена особая структура - пиреноид. Это - белковое образование, состоящее главным образом из рибулозобисфосфаткарбоксилазы - фермента, который осуществляет фиксацию двуокиси углерода. Пиреноид участвует в запасании углеводов, например крахмала. Красный глазок воспринимает изменения в интенсивности освещения, и клетка либо перемещается туда, где интенсивность света оптимальна для фотосинтеза, либо остается на месте, если освещенность достаточна. Такая ответная реакция на свет называется фототаксисом (разд. 15.1.2). Клетка хламидомонады передвигается за счет биения двух жгутиков и ввинчивается в воду, как штопор, вращаясь вокруг продольной оси.
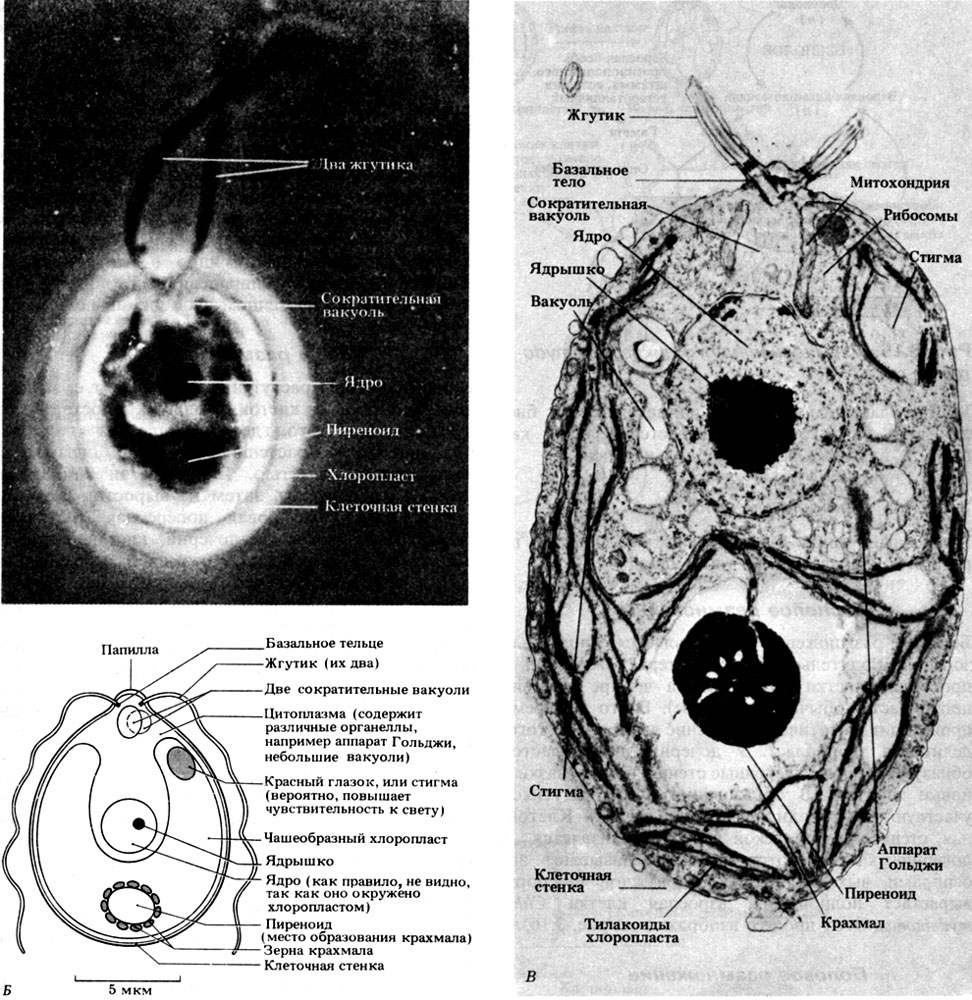
Рис. 3.14. A. Chlamydomonas в световом микроскопе; х 600. Б. Схема строения Chlamydomonas. В. Электронная микрофотография Chamydomonas reinhardtii. × 1400
Жизненный цикл
Жизненный цикл Chlomydomonas изображен на рис. 3.15. Взрослая особь гаплоидна.
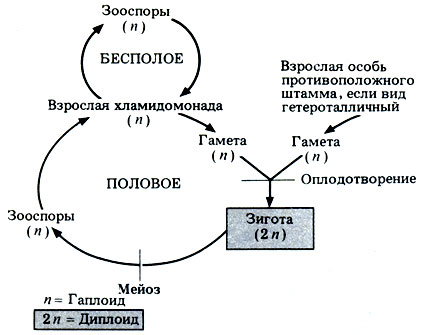
Рис. 3.15. Схема жизненного цикла Chlamydomonas
Бесполое размножение
Бесполое размножение осуществляется с помощью зооспор. Родительская клетка теряет жгутики, и протопласт клетки делится на два-четыре дочерних протопласта (обычно на четыре). В это же время происходит митотическое деление ядра; кроме того, делится и хлоропласт. У дочерних протопластов образуются новые клеточные стенки, новые глазки и новые жгутики. В образовании новых жгутиков участвуют центриоли (базальные тельца). Клеточная стенка родительской клетки ослизняется, и дочерние клетки, которые теперь называют зооспорами, выходят наружу. Из каждой зооспоры вырастает полноценная взрослая клетка Chlamydomonas. Этот процесс изображен на рис. 3.16, А.
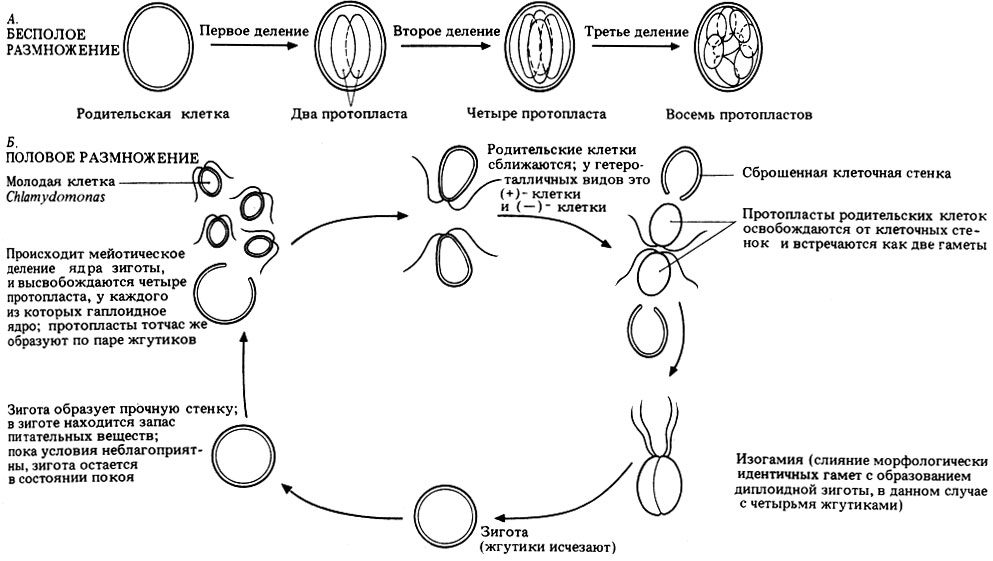
Рис. 3.16. Размножение Chlamydomonas, А. Бесполое размножение. Б. Половое размножение у изогамных видов
Половое размножение
Одни виды Chlomydomonas - гомоталличны, другие - гетероталличны; при этом разные виды могут быть изогамными, анизогамными или оогамными. Размножение изогамных видов изображено на рис. 3.16, Б. При прорастании ядро зиготы первый раз делится мейотически, при этом восстанавливается гаплоидное состояние, свойственное взрослым организмам. Высвободившиеся молодые клетки Chlomydomonas можно называть зооспорами, пока они полностью не созреют.
В прудах и других водоемах с непроточной, но чистой водой живет еще одна водоросль - неветвящаяся нитчатая водоросль Spirogyra. Большая часть видов спирогиры - плавающие формы, а нити у нее слизистые и скользкие.
Строение
Цилиндрические клетки спирогиры соединены торец в торец и образуют нить, изображенную на рис. 3.17. Все клетки идентичны, и между ними не наблюдается разделения функций. Тонкий слой цитоплазмы лежит по периферии клетки, а большая вакуоль как бы обмотана тяжами цитоплазмы. Такие тяжи удерживают ядро в центре клетки. Один или несколько спиралевидных хлоропластов лежат в тонком постенном слое цитоплазмы.
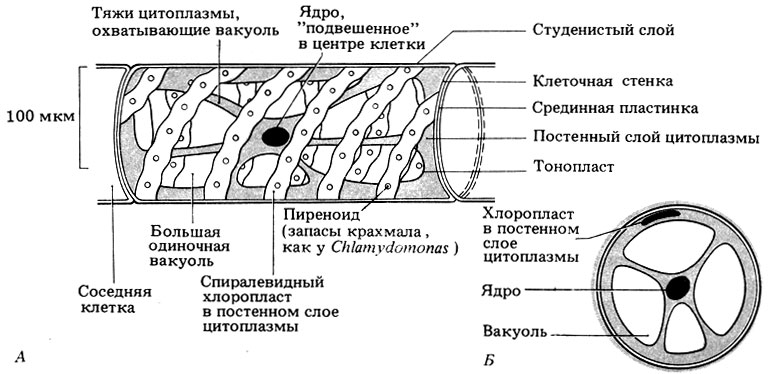
Рис. 3.17. Строение Spirogyra. А. Схематический вид сбоку. Б. Схема поперечного среза клетки в области ядра. Видно цилиндрическое строение клетки
Рост и размножение
Нити спирогиры растут интеркалярно, т. е. за счет деления любой из клеток, входящих в состав нити, независимо от того, где находится эта клетка. У большинства же растений зона роста ограничена верхушечной областью. Ядро клетки спирогиры делится митотически, затем из выростов боковых стенок образуется новая поперечная клеточная стенка. Получаются две дочерние клетки, которые вырастают до нормальных размеров, в результате чего вся нить увеличивается в длину.
Как мы уже отмечали (разд. 3.2.1), бесполое размножение происходит путем фрагментации.
Половое размножение осуществляется весьма специфическим способом, характерным для нитчатых водорослей: две нити располагаются бок о бок и супротивные клетки обеих нитей соединяются короткими трубчатыми выростами. Все содержимое клетки ведет себя как гамета; процесс этот можно рассматривать как анизогамный, поскольку, хотя обе гаметы и идентичны морфологически, только одна из них подвижна и перетекает в другую клетку через соединительную трубку. Такой процесс называется конъюгацией.
3.2.5. Отдел Phaeophyta
Основные признаки Phaeophyta перечислены в табл. 3.4.
У скалистых берегов Британского побережья часто встречаются разные водоросли из рода Fucus. Они очень хорошо приспособились к достаточно суровым условиям литоральной зоны, т. е. той зоны, которая попеременно то обнажается при отливе, то снова покрывается водой.
Наиболее известны три вида Fucus, которые чаще других встречаются у побережья в трех разных зонах на разной глубине; такое явление называется зональным распределением. Эти водоросли распределяются по зонам в соответствии с их способностью выдерживать пребывание на воздухе. Перечислим основные признаки, по которым их можно узнать, и места на берегу, где их можно найти:
F. spiralis (эти плоские водоросли выбрасывает на берег море) - у высшей точки прилива. В погруженном состоянии таллом слегка закручен в спираль.
F. serratus (то, что называют обыкновенными, зубчатыми или пильчатыми водорослями) - в средней приливной зоне. Края таллома зазубрены.
F. vesiculosus (так называемые пузырчатые водоросли) - у высшей точки отлива. Имеются воздушные пузыри, которые обусловливают плавучесть. На рис. 3.18 можно видеть характерные внешние признаки F. vesiculosus, а на рис. 3.19 изображены основные особенности его внутреннего строения.
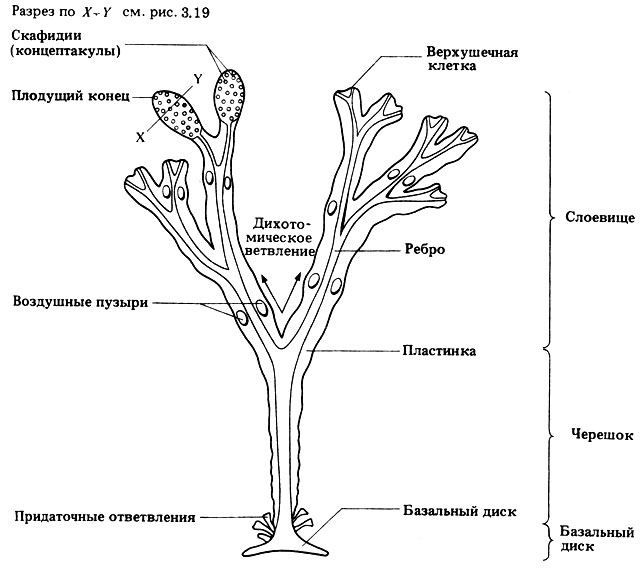
Рис. 3.18. Внешнее строение Fucus vesiculosus. Отмечены характерные признаки и, в частности, приспособления к окружающей среде. Плодущий конец (рецептакул) представляет собой набухшую и покрытую мелкими вздутиями (скафидиями или концептакулами), сообщающимися с наружной средой только узкими отверстиями, часть таллома. У женских растений плодущие концы темно-зеленые, у мужских оранжевые. Воздушные пузыри обычно парные и придают водоросли плавучесть. Придаточные ответвления (иногда отламываются; это одна из форм вегетативного размножения). Верхушечная клетка представляет собой точку роста, где происходит деление клеток. Ребро - это жесткое образование, которое выполняет механические функции и, возможно, участвует в переносе некоторых веществ. Пластинка плоская и упругая (кожистая); зеленовато-коричневого цвета из-за близкого к поверхности фотосинтезирующего слоя; покрыта слизью, предохраняющей от высыхания при отливе. Ребро вместе с пластинкой образуют таллом. Укореняющаяся часть таллома (в данном случае базальный диск) бесцветна и очень прочно прикрепляет таллом к скалам и т. п. Размеры водоросли варьируют в пределах до 1 м или более. Слоевище плоское и ремневидное; характер ветвления таков, что сопротивление волнам сводится к минимуму; воздушные пузыри поддерживают слоевище у поверхности, что способствует фотосинтезу. Черешок - это в основном ребро; черешок гибкий и потому успешно противостоит волнам
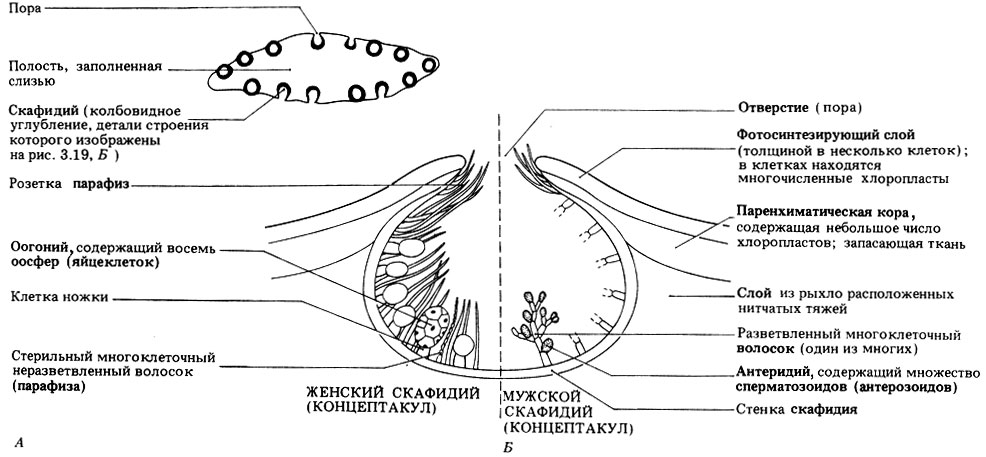
Рис. 3.19. Органы размножения Fucus vesiculosus. А. Вертикальный разрез плодущего конца (при малом увеличении). Б. Вертикальный разрез скафидия (при большом увеличении)
В теле водоросли, или талломе, наблюдается некоторое разделение функций между разными тканями. Эта тенденция у Phaeophyta прослеживается лучше, чем у всех остальных групп водорослей. Приспособления водорослей к окружающей среде мы рассмотрим чуть позже.
Органы размножения
Половое размножение оогамное. F. vesiculosus и F. serratus - двудомные растения, т. е. у них есть и мужские, и женские особи. F. spiralis - гермафродит, у которого на одном растении в одних и тех же вместилищах - скафидиях, или концептакулах, - находятся и мужские, и женские репродуктивные органы. Репродуктивные органы развиваются внутри скафидиев на "плодущих" кончиках некоторых слоевищ. В каждом скафидии имеется узкое отверстие (пора), через которое впоследствии высвобождаются наружу репродуктивные органы. Их строение показано на рис. 3.19.
Взрослые растения диплоидные, а гаметы образуются в результате мейотического деления.
Приспособления к окружающей среде
Прежде чем мы рассмотрим приспособления Fucus к среде обитания, следует сказать несколько слов о самой среде, которая достаточно враждебна. Будучи растениями приливно-отливной зоны, разные водоросли в разной степени подвергаются воздействию воздушной среды во время отлива. Поэтому у них должны быть защитные приспособления от высыхания. К тому же и температура очень резко меняется, когда холодные морские волны вливаются в прогретые лужицы, оставшиеся после отлива. Растения должны быть приспособлены и еще к одному фактору, а именно к резким изменениям солености воды, будь то ее увеличение при испарении из небольших водоемов, образовавшихся после отлива, или ее уменьшение во время дождя. Для того чтобы противостоять таким факторам, как приливы, отливы, прибой и удары волн, нужна достаточная механическая прочность. Большие волны начинают перекатывать камни, и это может очень сильно повредить растения.
Морфологические приспособления (общее строение)
Слоевище водоросли прочно прикреплено к грунту укореняющейся частью таллома (ризоидами или базальным диском) (рис. 3.18). Оно настолько прочно связывается с грунтом (обычно это камни), что водоросль чрезвычайно трудно оторвать от него. Как правило, первым не выдерживает камень, а не укореняющаяся часть таллома.
Таллом водорослей не сплошной, а рассеченный; он дихотомически ветвится в одной плоскости, и это позволяет свести к минимуму сопротивление толще воды. К тому же он прочный и упругий, но не жесткий. Ребра слоевища крепкие и гибкие.
У плавучей водоросли F. vesiculosus имеются специальные воздушные пузыри, которые удерживают слоевище у поверхности воды, т. е. в условиях, способствующих максимальному улавливанию света для фотосинтеза.
Физиологические приспособления
Среди фотосинтетических пигментов преобладает бурый пигмент - фукоксантин. Это - одно из приспособлений к фотосинтезу под водой, так как фукоксантин сильно поглощает синий свет, который проникает в толщу воды гораздо дальше, чем более длинноволновые лучи, например красные.
Таллом выделяет много слизи, которая заполняет все внутренние полости водоросли и просачивается наружу. Слизь помогает лучше удержать воду и препятствует обезвоживанию.
Осмотическое давление в клетках намного выше, чем в морской воде, поэтому осмотических потерь воды не наблюдается.
Приспособления к половому размножению
Выход гамет синхронизирован с приливами. Во время отлива таллом обсыхает, и из скафидиев наружу выдавливаются репродуктивные органы, которые от высыхания предохраняются слизью. Во время прилива стенки репродуктивных органов растворяются, высвобождая гаметы. Мужские гаметы подвижны и обладают положительным хемотаксисом в отношении веществ, выделяемых женскими гаметами.
Развитие зиготы происходит сразу же после оплодотворения, что сводит к минимуму риск быть унесенной в океан.
3.2.6. Отдел Euglenophyta
Основные признаки Euglenophyta приведены в табл. 3.4. Для этого отдела характерны признаки как растений, так и животных, что сильно затрудняет классификацию относящихся сюда организмов. По этой причине их обычно включают в свои систематические схемы и ботаники, и зоологи. Эти проблемы мы обсудим позднее, после описания рода Euglena.
Euglena - самая обычная одноклеточная водоросль, живущая в пресноводных прудах, канавах и любых других водоемах, богатых растворенными органическими соединениями. Как и Chlamydomonas, она иногда размножается так интенсивно, что вода становится зеленой, потому что среди пигментов эвглены преобладает хлорофилл. Строение эвглены показано на рис. 3.20, где отмечены и некоторые ее особенности.
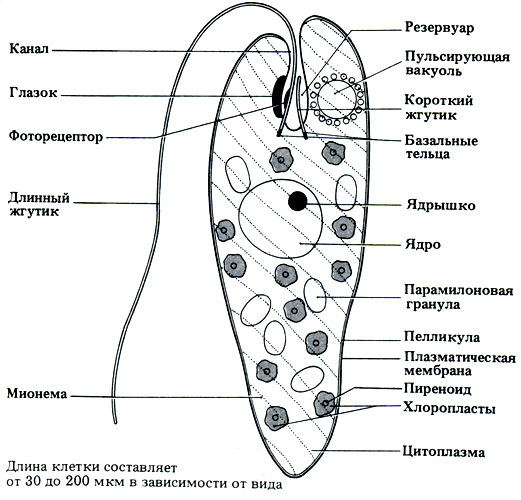
Рис. 3.20. Строение Euglena gracillis. Канал - место, через которое поступает пища у незеленых видов; пелликула здесь отсутствует, что позволяет заглатывать мелкие частички. Глазок (стигма) имеет красный цвет; участвует в реакции фототаксиса. Фоторецептор обнаруживает источник света и заставляет организм плыть в направлении оптимальной освещенности (фототаксис); направление движения может меняться при затенении фоторецептора. Длинный жгутик используется для локомоции; обычно направлен вперед; волнообразные движения проходят по жгутику от основания к кончику; жгутик тащит за собой клетку; во время движения вперед клетка вращается вокруг своей оси, оставляя за собой штопорообразный след. Пульсирующая вакуоль окружена вспомогательными вакуолями; участвует в осморегуляции, выкачивая в резервуар избыток воды, поступившей в клетку в результате осмоса. Короткий жгутик не участвует в локомоции. Парамилоновая гранула образована полимером глюкозы, похожим на крахмал и являющимся запасным углеводом. Пелликула располагается под плазматической мембраной; гибкая. Хлоропласты содержат фотосинтетические пигменты. В цитоплазме находятся сократительные волокна, которые обусловливают перистальтические волны деформации клетки; такое движение называется эвгленоидным
У Euglena нет клеточной стенки. Снаружи клетка покрыта плазматической мембраной, сразу же под которой находится белковая пелликула. Пелликула довольно гибкая, и это позволяет клетке принимать разную форму. Пелликула полностью окружает цитоплазму, и ее можно рассматривать как своего рода наружный скелет. Она состоит из ряда утолщенных продольных полосок и микрофибрилл, переплетенных между собой. Когда внутри цитоплазмы сокращаются крошечные фибриллы, которые называются мионемами, полоски пелликулы начинают скользить относительно друг друга, в результате чего изменяется форма тела. Это явление называется эвгленоидным движением. Другой, более обычный для эвглены способ передвижения за счет вращения длинного жгутика изображен на рис. 3.20 (рассмотрите глазок, фоторецептор и длинный жгутик) и подробно описан в разд. 17.6.3.
Бесполое размножение происходит посредством продольного деления клетки надвое. Полового размножения не наблюдается.
Питание
Зеленые виды Euglena автотрофны и синтезируют все необходимые им вещества из двуокиси углерода, воды и минеральных солей. Вместе с тем они нуждаются в поступлении извне витаминов В1 и В12, которые они не могут синтезировать сами. В этом Euglena не отличается от животных, хотя такая потребность в витаминах характерна и для многих других водорослей.
У нескольких видов Euglena хлорофилла нет, и поэтому они не окрашены и не способны к фотосинтезу (т. е. гетеротрофны). Питаются они по типу сапрофитов, переваривание происходит вне клетки. Когда водоем загрязнен, они процветают, так как разлагающийся материал богат органическими соединениями. Другие бесцветные формы способны заглатывать мелкие частички пищи, для чего у них имеется своеобразная "глотка", где отсутствует пелликула. Затем эти частички перевариваются внутри клетки (голозойное питание, разд. 10.1.1). Пища загоняется в глотку за счет движения жгутиков. Эти виды во многом напоминают простейшее Реrапеmа (разд. 4.1.1).
Если зеленые клетки Euglena долго держать в темноте, то хлоропласты исчезают, и клетки становятся бесцветными. Если в среде достаточно органических веществ, то клетки могут долго жить как сапрофиты. Когда их переносят на свет, снова появляется хлорофилл.
Проблемы систематики Euglena
Как мы уже говорили и как это следует из табл. 3.1, для Euglena характерны признаки и растений, и животных. Один из таких животных признаков, который мы еще не рассматривали, - это наличие в глазке астаксантина - пигмента, свойственного животным.
Легкость, с которой некоторые эвглены могут переходить из зеленой формы в бесцветную и наоборот, свидетельствует о том, что постоянно бесцветные виды, по-видимому, произошли от зеленых. Если впоследствии у бесцветных форм возникли специальные приспособления для голозойного питания, подобные тем, которые имеются у Peranema, то вполне возможно, что предки простейших были похожи на растения. Не следует забывать, однако, что эволюция могла идти и в обратном направлении, ведь мы уже обсуждали в начале этой главы возможность того, что предки растений могли быть похожи на животных (т. е. на гетеротрофных эукариот).
Решая вопрос о том, в царство растений или в царство животных помещать Euglena, необходимо помнить, что некоторые признаки животных имеются и у хламидомонады, и тем не менее ее обычно относят к растениям. Основные затруднения систематиков связаны со способом питания. Судя по всему, эвглену все же следует относить к растениям, так как наличие хлоропластов считается уникальной особенностью, присущей только царству растений. Все это, однако, лишний раз напоминает нам, сколь трудно навязать природе искусственную систематику, придуманную людьми.
| Среди растений | Среди животных | ||||
| Царство: | Plantae | Царство: | Animalia | ||
| Отдел: | Euglenophyta | Тип: | Protozoa | ||
| Класс: | Euglenophyceae | Класс: | (Flagellata) Mastigophora | ||
| Род: | Euglena | Род: | Euglena | ||
3.3. Составьте таблицу растительных и животных признаков Euglena. Воспользуйтесь для этого табл. 3.1, рис. 3.20 и сведениями, изложенными выше.
3.2.7. Направления эволюции водорослей
Даже тех нескольких примеров, которые мы рассмотрели в предыдущих разделах, вполне достаточно, чтобы понять, что существует множество типов водорослей, включая и такие одноклеточные формы, как Chlamydomonas, и такие сравнительно крупные организмы, как Fucus, у которых тело дифференцировано и наблюдается определенное разделение функций между отдельными тканями. У некоторых крупных бурых водорослей имеются даже проводящие ткани, хотя настоящей проводящей ткани - ксилемы и флоэмы - у них нет.
У водорослей четко прослеживается тенденция к усложнению процесса полового размножения от простой изогамии и анизогамии к оогамии. Однако следует с большой долей осторожности использовать ту или иную тенденцию для объяснения эволюционных взаимосвязей между отдельными группами водорослей. Такие взаимосвязи до сих пор окончательно не выяснены, а группа Chlorophyta (зеленые водоросли), от которой, как полагают, произошли наземные растения, отличается очень большим разнообразием: в ней есть и простые одноклеточные формы, и гораздо более сложные, а половое размножение также варьирует от изогамии до оогамии.
3.2.8. Значение водорослей
Роль водорослей в биосфере
По современным оценкам, на долю океана приходится по меньшей мере половина мировой первичной продукции, выражающейся в количестве фиксированного углерода. Эту первичную продукцию образуют водоросли - единственные растения, которые населяют океан. Учитывая ту огромную площадь, которую занимает океан, следует ожидать, что его продуктивность должна быть еще больше, но нельзя забывать, что фотосинтез возможен только в поверхностных слоях, куда проникает свет и где лимитирующим фактором является доступность биогенных элементов, особенно азота и фосфора.
Водоросли - очень важные первичные продуценты (гл. 12), с которых начинается большинство пищевых цепей, в том числе практически все морские и многие пресноводные цепи. Эти цепи через зоопланктон*, ракообразных и т. п. доходят до рыб. Многие микроскопические водоросли - одноклеточные, и именно они являются главным компонентом фитопланктона*.
* (Планктон это мельчайшие растения (фитопланктон) и животные (зоопланктон), которые свободно плавают в поверхностных слоях океанов и озер. Планктон имеет очень большое хозяйственное и экологическое значение.)
Фиксация углерода - это только одно из следствий фотосинтеза (разд. 9.2). Кроме того, благодаря фотосинтезу поддерживается уровень кислорода в атмосфере, при этом по меньшей мере половину всего кислорода выделяют водоросли, и их вклад в этот процесс намного больше, чем вклад наземных лесов.
Альгиновая кислота, агар и каррагенан
Из водорослей получают многие полезные продукты, например альгиновую кислоту, агар и каррагенан. Альгиновая кислота и ее производные (альгинаты) - это полисахариды, которые экстрагируют из срединной пластинки и клеточных стенок таких бурых водорослей, как Laminaria, Ascophyllum и Macrocystis. Водоросли добывают в большом количестве в прибрежных мелких водах; Macrocystis, например, собирают на побережье Калифорнии. Очищенные альгинаты не токсичны и легко образуют гели. Их широко применяют в качестве отвердителей и желеобразующих веществ для получения промышленных товаров (например, в косметике - для изготовления кремов для рук); в качестве эмульгаторов - для приготовления мороженого; в качестве желеобразующих веществ - в кондитерской промышленности; при изготовлении лаков, красок и лекарств; для получения глазурованной керамической посуды.
Агар - полисахарид, который получают из красных водорослей. Он образует такие же гели, как и альгинаты, но, возможно, более известен, так как является очень удобной средой для выращивания бактерий и грибов. С этой целью готовят разбавленный раствор агара, затем добавляют в него различные питательные вещества, стерилизуют и дают застыть, получая желеобразную массу. Кроме того, агар используют для тех же самых целей, что и альгинаты.
Каррагенан (карраген) - это еще один полисахарид клеточной стенки, который получают главным образом из красной водоросли Chondrus crispus. По своей химической структуре он очень похож на агар и применяется для тех же самых целей.
Диатомит (кизельгур)
Водоросли, относящиеся к отделу Bacillariophyta, в основном одноклеточные; их называют диатомовыми. Для этих водорослей характерно особое строение клеточной стенки, в которой содержится кремний. После гибели клеток остатки диатомей падают на дно морей и озер, и постепенно там накапливаются большие отложения. Образующаяся таким образом "диатомовая земля" содержит очень много (до 90%) кремния. После соответствующей очистки эту "землю" можно использовать как превосходный фильтрующий материал (например, при получении сахара или для осветления пива), как наполнитель при изготовлении красок или бумаги и как изоляционный материал, способный противостоять резким перепадам температуры.
Удобрение
На фермах, расположенных вблизи побережья, крупные водоросли (красные и бурые) по традиции используют как удобрения, хотя и в незначительных масштабах. Водоросли богаты калием, но в них гораздо меньше азота и фосфора, чем в простом навозе. Поэтому их удобряющее действие не очень велико. Более значимую роль играют свободноживущие сине-зеленые водоросли, которые являются очень важными азотфиксаторами и довольно распространены в почве (разд. 9.11.1).
Пищевые продукты
Некоторые водоросли подают прямо к столу, особенно на Дальнем Востоке. Красную водоросль Porphyra, считающуюся деликатесом, и большую бурую водоросль Laminaria обычно едят в сыром виде или же готовят из них разные блюда. В Южном Уэльсе Porphyra кладут в одно из традиционных блюд, для приготовления которого отваренные водоросли смешивают с овсянкой и потом все это тушат в масле. В поисках новых источников пищи много внимания было уделено промышленному культивированию водорослей. Однако для получения новых пищевых продуктов годятся очень немногие водоросли, и до сего времени сколько-нибудь значимые успехи в этой области были достигнуты при культивировании бактерий и грибов. Из сине-зеленых водорослей многообещающей считается Spirulina.
Очистка сточных вод
Водоросли вносят определенный вклад в работу микроорганизмов по очистке сточных вод, так как в сточных водах содержатся питательные вещества не только для бактерий, грибов и простейших, но и для микроскопических зеленых водорослей. Они особенно полезны в открытых "окислительных прудах", которые достаточно широко используются в тропических и субтропических странах. Открытые пруды глубиной от 1 до 1,5 м заливают неочищенными стоками. В процессе фотосинтеза водоросли выделяют кислород и обеспечивают таким образом жизнедеятельность других аэробных микроорганизмов, растущих в сточных водах. Время от времени водоросли собирают и перерабатывают на корм скоту.
Научные исследования
Одноклеточным водорослям присущи все характерные признаки типичных растений, поэтому они являются идеальным материалом для научных исследований, так как, во-первых, их можно выращивать в большом количестве в строго определенных условиях и, во-вторых, для этого не требуется очень много места. Примером таких водорослей может служить Chlorella, которой по праву принадлежит почетное место в исследованиях фотосинтеза (разд. 9.4.3). Водоросли используются и при изучении поглощения ионов. Они принесли большую пользу и в новаторских исследованиях строения клеточной стенки и жгутиков.
Вред, наносимый водорослями
В определенных условиях водоросли "цветут", т. е. в огромных количествах скапливаются в воде. "Цветение" наблюдается при достаточно теплой погоде, когда в воде много питательных веществ. Такая ситуация очень часто искусственно создается человеком, когда в воду сбрасывают промышленные стоки или же когда в реки и озера попадают удобрения с полей. В результате начинается взрывоподобное размножение первичных продуцентов (водорослей), и они в нарушение всех законов природы начинают отмирать раньше, чем их успеют съесть. При последующем разложении остатков происходит столь же интенсивное размножение аэробных бактерий и вода полностью лишается кислорода. Все это происходит очень быстро, и из-за нехватки кислорода начинают гибнуть рыбы и другие животные и растения. Увеличение концентрации питательных веществ в воде, которое запускает весь этот процесс, называется эвтрофизацией водоема, и если оно происходит быстро, то можно считать, что это еще одна из форм загрязнения окружающей среды.
Токсины, образующиеся при "цветении" воды, в особенности при размножении сине-зеленых водорослей, увеличивают гибель животных. Подобные взрывы численности водорослей представляют собой серьезную проблему для рыбоводческих хозяйств, особенно там, где интенсивный вывоз удобрений на поля еще в большей степени усиливает эвтрофизацию. Сходные осложнения возникают и при "цветении" воды в океане. Кроме того, токсины, накапливаясь в теле моллюсков и ракообразных, питающихся водорослями, и затем попадая в организм человека, вызывают у него различные отравления и паралич.
С водорослями связаны и многие сложности при хранении питьевой воды в запасных резервуарах, когда она загрязняется продуктами жизнедеятельности водорослей или когда водоросли начинают расти на песчаных фильтрах, полностью забивая их.
3.4. Те трудности, о которых мы только что говорили, чаще возникают в водохранилищах, расположенных в низинах. Объясните, почему это так и должно быть.
3.5. В отличие от многих грибов и бактерий водоросли не вызывают никаких заболеваний. С чем это связано?
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'