3.5. Отдел Spermatophyta - семенные растения
Spermatophyta (греч. sperma - семя) - самая процветающая группа наземных растений. В этом разделе основное внимание мы уделим тем адаптационным приспособлениям семенных растений, которые способствовали их процветанию, и, кроме того, сравним их с более низко организованными группами, которые мы уже рассмотрели.
Семенные растения, по-видимому, произошли от вымерших семенных папоротников. Если вспомнить Selaginella (как одного из представителей папоротникообразных), то следует отметить, что у нее, по существу, такой же жизненный цикл, как и у семенных растений; разница лишь в том, что у Selaginella женский гаметофит автотрофен, а у семенных растений он утрачивает автотрофность. Однако давайте забудем о селагинелле и попытаемся сравнить жизненный цикл семенных растений и равноспоровых папоротникообразных (обыкновенных папоротников).
Одна из основных трудностей, с которой сталкиваются растения на суше, связана с уязвимостью гаметофитного поколения. Так, например, у папоротников гаметофит - это нежный заросток, который образует мужские гаметы (сперматозоиды), нуждающиеся в воде, чтобы достичь яйцеклетки. А у семенных растений гаметофит защищен и очень сильно редуцирован. Только сравнив жизненные циклы семенных и более примитивных растений, можно понять, что и у семенных растений сохраняется чередование поколений. Семенные растения обладают тремя очень важными преимуществами: 1) разноспоровостью, 2) образованием семян и 3) появлением неплавающих мужских гамет.
Разноспоровость
Очень важным шагом на пути эволюции от папоротникообразных к семенным растениям было появление растений, образующих споры двух типов - микроспоры и мегаспоры. Такие растения называют разноспоровыми; они были рассмотрены в разд. 3.4. В табл. 3.6 приведен краткий словарь терминов, относящихся к спорообразованию в жизненном цикле разноспоровых растений (см. также рис. 3.26). Все семенные растения разноспоровые.
Из микроспоры развивается мужской гаметофит, а из мегаспоры возникает женский гаметофит. И в том и в другом случае гаметофит очень сильно редуцирован и не выходит из споры. Исключение составляет свободно живущий самостоятельный гаметофит равноспоровых растений, таких, как Dryopteris. Спора защищает гаметофит от высыхания, что является важным приспособлением к жизни на суше. Гаметофиты не способны к фотосинтезу, поэтому они нуждаются в запасах питательных веществ, накопленных в спорах предшествующим спорофитным поколением. Как мы увидим дальше, предельная редукция гаметофита наблюдается у цветковых растений.
Мегаспоры образуются в мегаспорангиях на мегаспорофиллах, а микроспоры - в микроспорангиях на микроспорофиллах. У семенных растений структура, эквивалентная мегаспорангию, называется семязачатком. Внутри семязачатка развивается всего одна мегаспора, или один женский гаметофит, который называется зародышевым мешком. Структура, эквивалентная микроспорангию, называется пыльцевым мешком. В пыльцевом мешке образуется много микроспор, которые называются пыльцевыми зернами или пылинками.
Эволюция семени
У семенных растений мегаспоры не отделяются от спорофита. В отличие от той картины, что мы наблюдаем у более примитивных разноспоровых организмов, мегаспоры остаются внутри семязачатков (мегаспорангиев), прикрепленных к спорофиту. Внутри мегаспоры развивается женский гаметофит (зародышевый мешок) и образуется одна или несколько женских гамет, или яйцеклеток. После оплодотворения женской гаметы семязачаток называют уже семенем. Таким образом, семя - это оплодотворенный семязачаток. Семязачаток, а позднее и семя обладает целым рядом преимуществ:
1. Женский гаметофит защищен семязачатком, целиком зависит от родительского спорофита, но гораздо менее чувствителен к обезвоживанию, чем свободно живущий гаметофит.
2. После оплодотворения образуется запас питательных веществ, получаемый гаметофитом от родительского спорофитного растения, от которого он по-прежнему не отделен. Этот запас используется развивающейся зиготой (следующим спорофитным поколением) после прорастания семени.
3. Семена приспособлены к тому, чтобы противостоять неблагоприятным условиям, и могут оставаться в состоянии покоя до тех пор, пока условия не станут благоприятными для прорастания.
4. У семян могут развиваться различные приспособления, облегчающие их распространение. Семя представляет собой сложную структуру, в которой собраны клетки трех поколений - родительского спорофита, женского гаметофита и зародыша следующего спорофитного поколения. В самом общем виде это изображено на рис. 3.34. Родительский спорофит дает семени все, что нужно для жизни, и только после того, как семя полностью созреет, т. е. накопит запас питательных веществ для зародыша спорофита, оно отделяется от родительского спорофита.
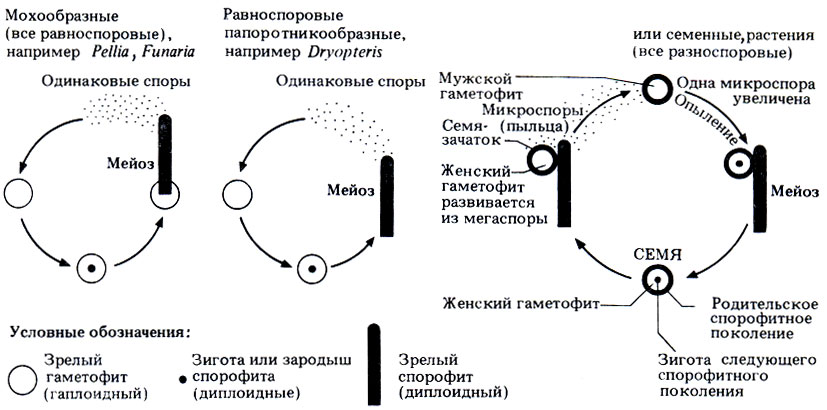
Рис. 3.34. Связь между гаметофитным и спорофитным поколениями у разных групп растений
Эволюция неплавающих мужских гамет и оплодотворения, независимого от воды
Для полового размножения растений, которые мы уже рассмотрели, необходимо, чтобы сперматозоид мог подплыть к яйцеклетке, т. е. нужна вода. Поэтому перед семенными растениями встают определенные проблемы. Для того чтобы произошло оплодотворение, мужские гаметы должны достичь женских гамет, а, как мы уже говорили, мужские и женские гаметы развиваются отдельно, и к тому же женские гаметы еще и остаются внутри семязачатков спорофита. Мужские гаметы образуются мужскими гаметофитами внутри микроспор, или пыльцевых зерен. Они не превращаются в плавающие сперматозоиды, а остаются неподвижными и переносятся вместе с пыльцевыми зернами из пыльцевых мешков (микроспорангиев) к семязачаткам. Такой перенос пыльцы называется опылением. На последнем этапе опыления образуется пыльцевая трубка, которая растет в сторону семязачатка; по этой трубке неподвижные мужские гаметы достигают яйцеклетки, и происходит оплодотворение. Вода не нужна спермиям ни на одной из перечисленных стадий. Только у некоторых примитивных семенных растений, таких, как саговниковые, спермин выходят из пыльцевых трубок, что указывает на определенную связь с несеменными растениями. На рис. 3.34 сравниваются жизненные циклы семенных и некоторых несеменных растений. Особо выделены происхождение семян и взаимосвязь между спорофитным и гаметофитным поколениями. Опыление, возможно, и не дает никакой выгоды, так как этот процесс носит чисто случайный характер и достигается с трудом, а образование большого количества пыльцы биологически невыгодно. Считают, что первоначально опыление происходило только с помощью ветра. Однако уже на заре эволюции цветковых растений появились первые летающие насекомые (около 300 млн. лет назад, в каменноугольном периоде). Сразу же возникла возможность более эффективного опыления насекомыми. Одна из групп семенных растений - цветковые растения - наиболее успешно использует эту возможность.
3.12. Шансы для выживания и возникновения пыльцевых зерен (микроспор), переносимых ветром, намного меньше, чем для спор Dryopteris. Почему?
3.13. Объясните, почему мегаспоры крупные, а микроспоры мелкие.
3.5.1. Основные признаки и систематика Spermatophyta
Основные признаки и систематика Spermatophyta представлены в табл. 3.8.
| Отдел Spermatophyta (семенные растения) | |
|
Общие признаки
Разноспоровые, т. е. имеют два типа спор: микроспоры и мегаспоры; микроспора = пыльцевое зерно, мегаспора = зародышевый мешок. Зародышевый мешок (мегаспора) остается целиком закрытым в семязачатке (мегаспорангии); семя - это оплодотворенный семязачаток. Спорофит доминирует; гаметофитное поколение крайне редуцировано. Для полового размножения не нужна вода, потому что мужские гаметы не способны плавать (исключение составляют некоторые наиболее примитивные представители); чтобы оплодотворить яйцеклетки, они проникают в завязь через пыльцевую трубку Сложные по строению проводящие ткани в корнях, стеблях и листьях. |
|
| Класс Gymnospermae (главным образом хвойные; кроме того, тиссовые, саговниковые, гинкговые и др.) | Класс Angiospermae (цветковые растения) |
| "Голые" семена: это означает, что семена лежат открыто, т. е. не спрятаны в завязи. | Семена спрятаны в завязи. |
| Обычно образуют шишки, на которых появляются спорангии и споры. | Образуют цветки, в которых развиваются спорангии и споры. |
| Плоды не образуются, потому что нет завязи. | После оплодотворения из завязи образуется плод. |
| В ксилеме нет сосудов - только трахеиды; во флоэме нет клеток-спутниц - только альбуминовые клетки (сходные по функции с клетками-спутницами, но отличающиеся от них по происхождению). | Ксилема состоит из сосудов; флоэма содержит клетки-спутницы. |
| Подклассы: двудольные и однодольные (см. табл. 3.9). | |
В этой таблице рассматриваются две группы семенных растений - голосеменные и покрытосеменные. Последнюю группу обычно называют цветковыми растениями. У голосеменных семязачаток, а потом и семена располагаются на поверхности особых листьев, которые называют мегаспорофиллами или семенными чешуйками. Эти чешуйки собраны в шишки. У покрытосеменных семена закрыты, что еще лучше предохраняет гаметофит и образующуюся затем зиготу. Структуры, в которые заключены семена, называются плодолистиками. Считают, что плодолистики эквивалентны мегаспорофиллам (листьям), сложенным так, что они закрывают семязачатки (мегаспорангии). Плодолистик может быть один или их может быть много.
Полое основание плодолистика, или группы слившихся вместе плодолистиков, называется завязью. В завязи находятся семязачатки. После оплодотворения завязь называют плодом, а семязачатки - семенами. Или плоды, или семена (иногда и то и другое) часто имеют различные приспособления для рассеивания.
На рис. 3.35 в виде простых схем изображены для сравнения различные спороносные структуры сосудистых растений. Сравнение поможет вам понять некоторые термины, которые были использованы при изложении этого материала.
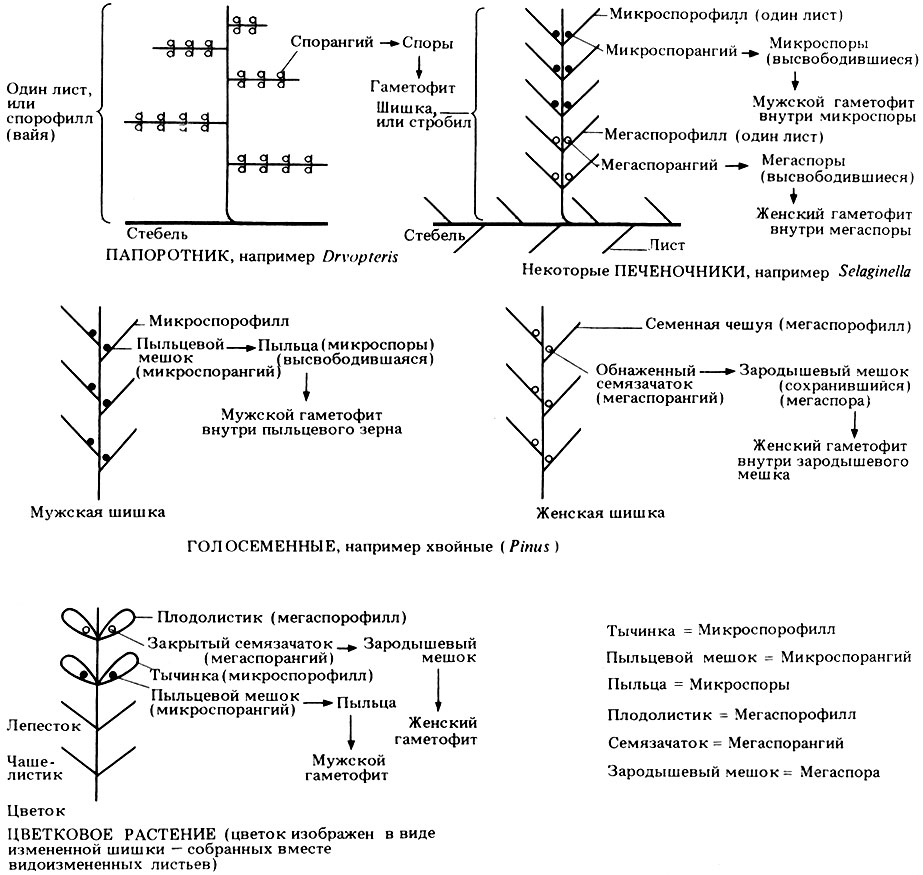
Рис. 3.35. Схематическое сравнение спороносных структур у сосудистых растений (папоротникообразных и семенных)
3.5.2. Класс Gymnospermae - голосеменные, например хвойные, саговниковые, тиссовые, гинкговые
Основные признаки Gymnospermae перечислены в табл. 3.8.
Голосеменные - процветающая группа растений, распространенных по всему земному шару; леса из представителей голосеменных составляют примерно треть всех лесов планеты. Среди голосеменных - деревья или кустарники, в основном вечнозеленые с листьями, похожими на иголки. Самая большая группа - хвойные, куда входят деревья, произрастающие в высоких широтах и распространяющиеся на север дальше всех других деревьев. Хвойные имеют большое хозяйственное значение, прежде всего как источник поделочной древесины, которая используется не только для получения пило- и лесоматериалов, но и для получения смолы, скипидара и древесной мезги. К хвойным относятся сосны, лиственницы (с опадающей на зиму хвоей), пихты, ели и кедры. Рассмотрим типичное хвойное дерево-сосну обыкновенную (Pinus sylvestris).
Pinus sylvestris распространена в Центральной и Северной Европе, СССР и Северной Америке. Она интродуцирована также в Великобританию, но в природных условиях растет только в Шотландии. Сосны выращивают и для декоративных целей, и для получения лесо- и пиломатериалов. Это - красивое величественное дерево высотой до 36 м с характерной отслаивающейся корой розовато- или желтовато-коричневого цвета. Сосны чаще всего растут на песчаных или небогатых горных почвах, и поэтому корневая система у них обычно расстилается по поверхности и сильно ветвится. Внешний вид сосны показан на рис. 3.36.
Каждый год из мутовки боковых почек на верхушке ствола вырастает новая мутовка ветвей. Характерный заостренный на конус облик Pinus и других хвойных обусловлен тем, что мутовки более молодых (и более коротких) веток на верхушке книзу постепенно сменяются все более старыми (и более длинными). С возрастом нижние ветки отмирают и отпадают, поэтому стволы старых деревьев обычно лишены ветвей (рис. 3.36).
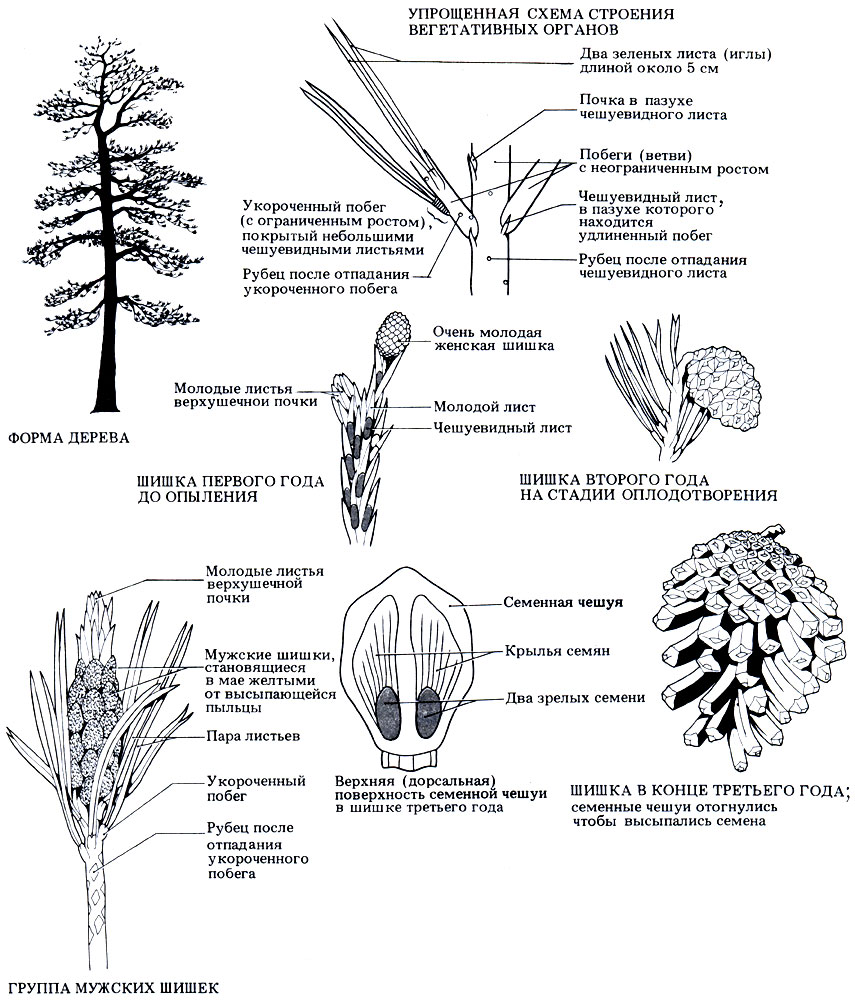
Рис. 3.36. Внешний вид и характерные особенности спорофитного поколения сосны обыкновенной (Pinus syIvestris)
Главные ветви и ствол продолжают расти из года в год за счет роста верхушечной почки. Поэтому говорят, что для хвойных характерен неограниченный рост. Чешуевидные листья расположены спиралью; в пазухах таких листьев находятся почки, из которых развиваются очень короткие веточки (длиной 2-3 мм), называемые укороченными побегами. Это - стебли с ограниченным ростом, на верхушке которых растет по два листа. Как только побег вырастет, чешуевидный лист у его основания отпадает, и на его месте остается рубец. Листья похожи на иголки, что уменьшает площадь поверхности листа, а следовательно, и потери воды. Кроме того, листья покрыты толстой восковой кутикулой, а устьица глубоко погружены в ткань листа - еще одно приспособление для сохранения воды. Ксероморфные приспособления вечнозеленых растений обеспечивают минимальную потерю воды во время холодных сезонов, когда вода замерзает, и ее трудно извлечь из почвы. Через два-три года укороченные побеги отпадают вместе с листьями, и на их месте остается еще один рубец.
Дерево представляет собой спорофит и является разноспоровым. Весной на дереве образуются и мужские, и женские шишки. Диаметр мужских шишек составляет около 0,5 см; они округлые и располагаются кучками у основания новых побегов под верхушечной почкой. Они образуются в пазухах чешуевидных листьев вместо укороченных побегов. Женские шишки появляются в пазухах чешуевидных листьев на конце новых сильных побегов на некотором расстоянии от мужских шишек и располагаются более беспорядочно. Полное развитие шишек занимает три года, поэтому все шишки имеют разные размеры, и на одном дереве можно обнаружить шишки от 0,5 до 6 см величиной. Молодые шишки зеленого цвета, а на второй год они становятся коричневыми или красновато-коричневыми. И мужские, и женские шишки состоят из плотно прижатых друг к другу спорофиллов, расположенных спиралью вокруг центральной оси (рис. 3.36).
На нижней поверхности каждого спорофилла мужской шишки находятся два микроспорангия, или пыльцевых мешка. Внутри пыльцевых мешков происходит мейотическое деление материнских клеток пыльцы и образуются пыльцевые зерна, или микроспоры. У пыльцевых зерен имеется по два воздушных мешка, которые помогают им переноситься ветром. В мае шишки становятся совсем желтыми из-за пыльцы, которая целым облаком вылетает из них. В конце лета они увядают и отпадают.
Спорофилл женской шишки состоит из нижней кроющей чешуйки и более крупной верхней чешуи, несущей семязачатки. На верхней поверхности крупной чешуи рядом находятся два семязачатка; в них происходит мейотическое деление материнской клетки мегаспоры и образуются четыре мегаспоры, из которых только одна развивается дальше. Опыление происходит еще на первом году развития шишки, но оплодотворение задерживается до следующей весны, когда прорастут пыльцевые трубки. Из оплодотворенных семязачатков образуются крылатые семена. Они продолжают зреть в течение второго года и высыпаются лишь на третий год. К этому времени шишка становится довольно крупной, одревесневает, а чешуи отгибаются наружу, перед тем как ветер разнесет семена.
3.5.3. Класс Angiospermae - покрытосеменные, или цветковые, растения
Основные признаки Angiospermae перечислены в табл. 3.8.
Покрытосеменные лучше других растений приспособлены к жизни на суше. Они появились в меловом периоде, около 135 млн. лет назад, быстро размножились, освоив самые разные местообитания, и вскоре вытеснили голосеменные, заняв главенствующее положение среди наземной растительности. Некоторые покрытосеменные вновь вернулись к пресноводному, а несколько видов - даже к солоноводному образу жизни.
Одна из самых характерных особенностей покрытосеменных, если не считать закрытые семена, о которых мы уже говорили, это - появление цветков вместо шишек. Наличие цветков позволило этим растениям привлечь для опыления насекомых, а иногда даже птиц и летучих мышей. Яркая окраска цветков, душистый аромат, съедобная пыльца и нектар-все это средства для привлечения животных. В некоторых случаях насекомые вообще не могут обходиться без цветков. Эволюция насекомых и цветков в ряде случаев была очень тесно связана, в результате чего возникли самые разные, очень специфические и притом взаимовыгодные отношения. Адаптация цветка, как правило, была направлена на максимальное увеличение шансов для переноса пыльцы насекомыми, и поэтому этот процесс более надежен, чем опыление ветром. Растениям, опыляемым насекомыми, не нужны такие большие количества пыльцы, как при опылении ветром. Тем не менее многие цветковые растения приспособились к опылению ветром.
Жизненный цикл
Жизненный цикл типичного цветкового растения изображен на рис. 3.37.
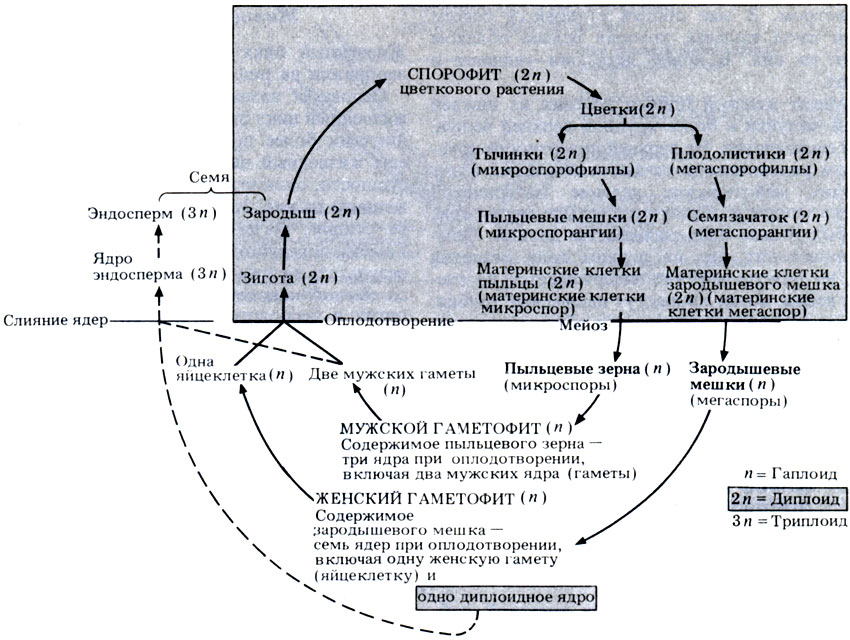
Рис. 3.37. Жизненный цикл покрытосеменных
Основное назначение этого рисунка - сравнить жизненный цикл цветкового растения с жизненными циклами более примитивных растений. Подробно сам жизненный цикл будет описан в разд. 20.2. В сущности, он почти не отличается от цикла, изображенного на рис. 3.21. Обратите особое внимание на те стадии, когда происходит мейоз или митоз. Гаметы образуются в результате митоза, а споры - в результате мейоза, как и у всех остальных растений со сменой поколений. Строго говоря, цветок - это орган и бесполого, и полового размножения, поскольку в нем образуются споры (бесполое размножение), внутри которых возникают гаметы (половое размножение). Следует отметить, что пыльцевое зерно является спорой, а не мужской гаметой, так как в нем самом находятся мужские гаметы. Как уже говорилось выше, пыльцевые зерна переносят мужские гаметы на женские репродуктивные органы, а это позволяет обойтись без плавающих сперматозоидов.
Процесс развития эндосперма также изображен на рис. 3.37. Из эндосперма образуются запасы питательных веществ, а сам способ их образования уникален и присущ только покрытосеменным.
Двудольные и однодольные
Покрытосеменные растения делятся на две большие группы, которым можно дать статус классов или подклассов в зависимости от того, какой систематической схемой пользоваться. Чаще всего эти две группы называют однодольными и двудольными. В табл. 3.9 перечислены основные признаки, по которым они различаются. Немногие из этих признаков в отдельности имеют систематическое значение, поскольку существуют многочисленные исключения, и лишь сочетание нескольких признаков позволяет точно идентифицировать такие растения. По современным представлениям однодольные - более совершенная группа; полагают, что они, вероятно, произошли от примитивных двудольных.

Таблица 3.9. Основные различия между двудольными и однодольными
Покрытосеменные растения бывают травянистые (т. е. не одревесневшие) и деревянистые. Деревянистые растения - это кустарники и деревья. У таких растений образуется большое количество вторичной ксилемы (древесины), которая служит внутренней опорой для ствола и, кроме того, выполняет функции проводящей ткани. Ксилема возникает в результате активности клеток камбия. Травянистые растения, или травы, полагаются только на тургесцентность клеток и на небольшое количество механических тканей, таких, как колленхима, склеренхима или ксилема; немудрено, что и сами они не очень велики. У травянистых растений либо совсем нет камбия, либо, если он и имеется, его активность незначительна. Многие травянистые растения однолетние, т. е. они за один год завершают свой цикл развития от семени до семени. У некоторых травянистых растений образуются многолетние органы типа луковиц, клубнелуковиц или клубней, которые перезимовывают или же переживают такие неблагоприятные условия, как засуха (разд. 20.1.1). В этом случае они бывают двулетними или многолетними, т. е. они либо образуют семена на второй год и отмирают, либо живут год за годом. Кустарники и деревья - многолетние растения и могут быть либо вечнозелеными, т. е. образуют и сбрасывают листву круглый год, и поэтому на растении всегда есть листья, либо листопадными, т. е. полностью сбрасывают листья в холодное или засушливое время. Чтобы проиллюстрировать, насколько разнообразны покрытосеменные, на рис. 3.38-3.42 показано строение некоторых представителей этого класса.

Рис. 3.38. Строение листа однодольных (А) и двудольных (Б) растений
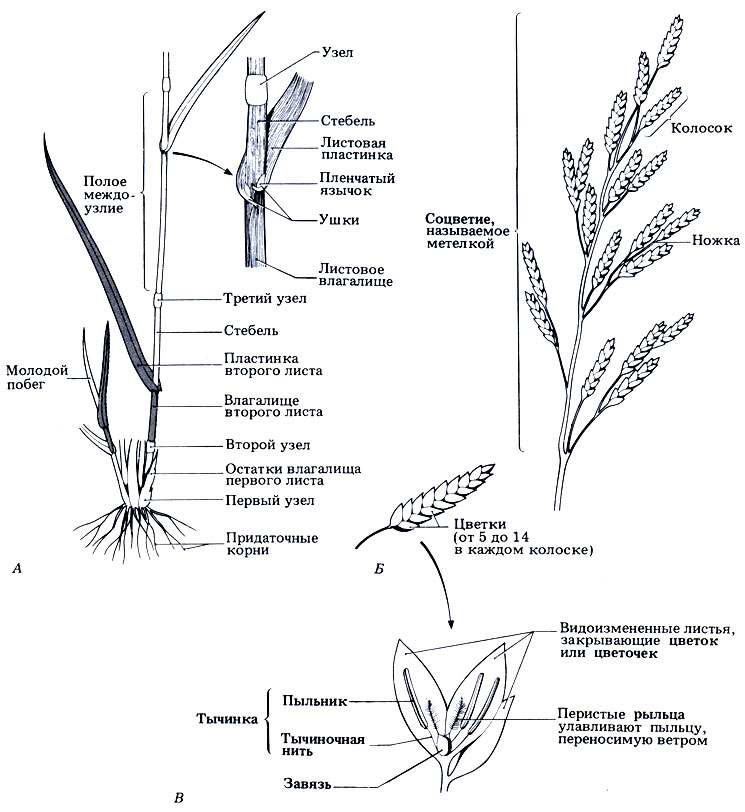
Рис. 3.39. Строение цветка и вегетативных органов однодольного травянистого растения - овсяницы луговой (Festuca pratensis). Это многолетнее растение высотой 30-120 см образует большие дерновины, встречается по всей Великобритании на заливных лугах, пастбищных угодьях, старых пастбищах и по обочинам дорог. Вторые листья на рисунке обозначены серым цветом. Листья, как правило, расположены двумя рядами попеременно то на одной, то на противоположной стороне стебля. А. Строение вегетативных органов. В узле имеется меристема, из которой вырастают лист и междоузлия; не полый в отличие от междоузлий. Для листовой пластинки характерно параллельное жилкование. Ушки представляют собой небольшие заостренные выступы (имеются не у всех злаков). Стебель неразветвленный; быстро удлиняется перед самым цветением, и тогда его называют соломиной. Влагалище второго листа цилиндрическое и частично закрывает междоузлие между вторым и третьим узлами. Придаточные корни отрастают от основания стеблей; образуют мочковатую корневую систему без стержневого корня. Молодой побег с еще не удлиненными междоузлиями; узлы расположены близко друг к другу и спрятаны во влагалища у основания побега. Стебель образуют узлы и междоузлия, а лист - листовая пластинка и влагалище. Б. Строение соцветия. В. Детали строения одиночного открытого цветка, или цветочка; не изображены две небольшие лепестковидные структуры (пленочки, или лодикулы), которые покрывают завязь
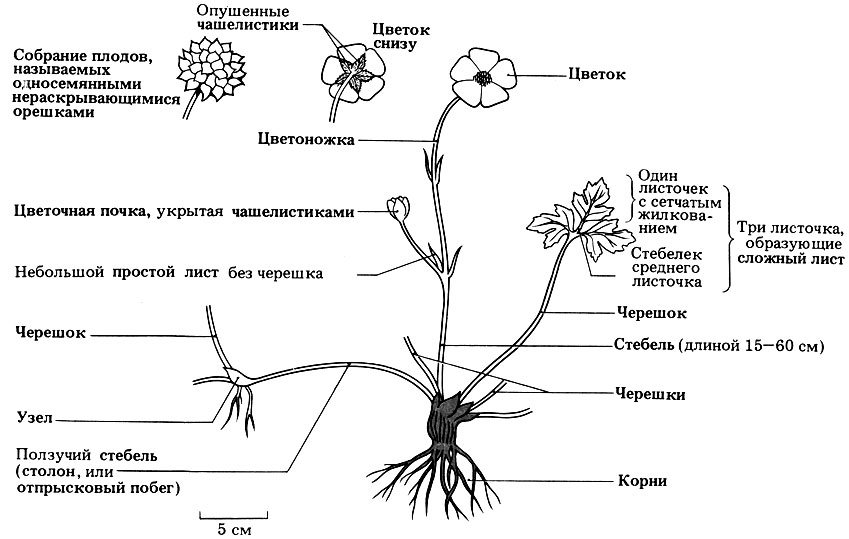
Рис. 3.40. Строение цветка и вегетативных органов двудольного травянистого растения - лютика ползучего (Ranunculus repens). Это обычное многолетнее растение встречается на мокрых лугах, в сырых лесах, садах и на брошенных пашнях по всей Великобритании. Цветки имеют пять раздельных блестящих лепестков золотисто-желтого цвета; тычинок и плодолистиков много; цветет с мая по август
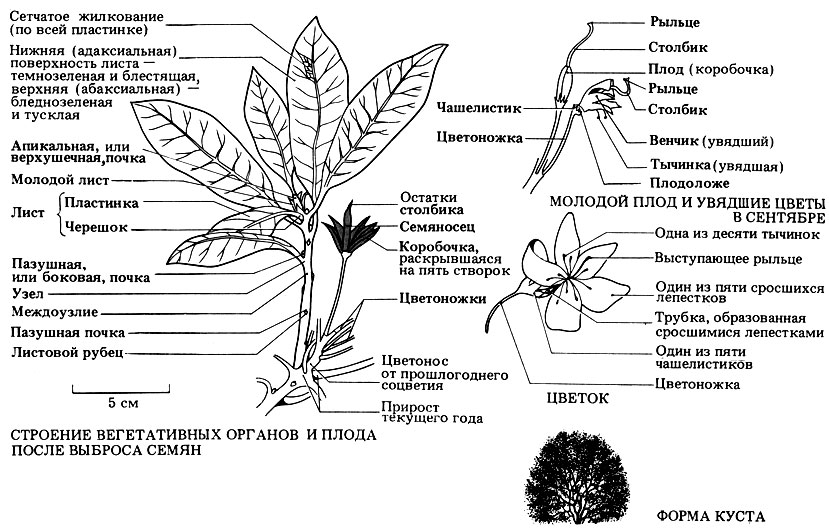
Рис. 3.41. Строение цветка и вегетативных органов дикого рододендрона (Rhododendron ponticum) - вечнозеленого двудольного кустарника, достигающего в высоту 3 м. Цветки рододендрона, появляющиеся в мае-июне, собраны в соцветия на конце побега; каждый цветок имеет довольно крупные размеры и по форме колоколообразный. Листья вдоль стебля располагаются поочередно, а вокруг верхушечной почки собраны в мутовку. Рододендроны обычно высаживают в парках и садах. Исходно интродуцированное, это растение успешно акклиматизировалось и хорошо растет на кислых почвах (песчаных или торфяных) на тех же местах, что и вереск, или в лесу
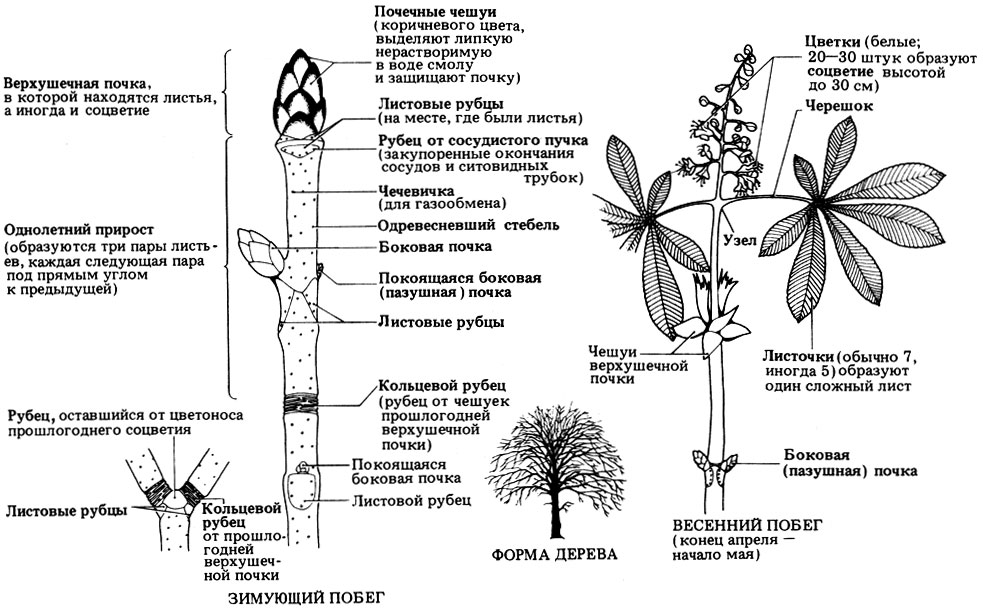
Рис. 3.42. Строение цветка и вегетативных органов конского каштана (Aesculus hippocastanum) - широколиственного двудольного дерева, теряющего листву на зиму. Высота дерева достигает 30 м и более
3.5.4. Краткое перечисление адаптационных приспособлений голосеменных и покрытосеменных растений к жизни на суше
Проблемы, связанные с переходом от водного образа жизни к наземному, мы уже затрагивали в разд. 3.3. Теперь, когда мы познакомились с представителями всех основных групп наземных растений, можно вновь вернуться к этому вопросу и обсудить, почему же голосеменные и покрытосеменные так хорошо приспособились к жизни на суше. Основное преимущество этих растений над всеми остальными, конечно же, связано с их способом размножения. Здесь можно выделить три главных аспекта:
1. Гаметофитное поколение очень редуцировано. Гаметофит полностью зависит от спорофита и всегда находится под его защитой. А у мхов и печеночников, у которых преобладает гаметофит, и у папоротников, у которых есть свободноживущий заросток, гаметофит не защищен и очень легко высыхает.
2. В отличие от всех других растений, у которых сперматозоиды подплывают к яйцеклеткам, покрытосеменным для оплодотворения не нужна вода. Мужские гаметы семенных растений неподвижны и переносятся ветром или насекомыми вместе с пыльцевыми зернами. На конечном этапе опыления мужские гаметы проникают к яйцеклетке через пыльцевую трубку, а сами яйцеклетки при этом заключены внутри семязачатков.
3. Из всех современных растений только у семенных есть особые структуры-семена. Возникновение семени стало возможным благодаря тому, что семязачатки вместе со всем своим содержимым остаются на родительском спорофите.
Другие характерные особенности покрытосеменных, помогающие им жить на суше, перечислены ниже. Подробнее мы обсудим их в соответствующих разделах этой книги.
а) У всех сосудистых растений ткани ксилемы и склеренхимы лигнифицированы и дают внутреннюю опору. У многих семенных растений наблюдаются вторичный рост и отложение большого количества древесины (вторичной ксилемы). К таким растениям относятся кустарники и деревья.
б) Настоящие корни, которые тоже характерны для сосудистых растений, позволяют эффективно извлекать влагу из почвы.
в) От высыхания эти растения защищены эпидермисом и нерастворимой в воде кутикулой или же пробкой, образующейся при вторичном утолщении.
г) Эпидермис наземных органов, и в особенности эпидермис листьев, пронизан устьицами, что способствует лучшему газообмену между растением и атмосферой. д) У растений есть и другие приспособления к жизни в жарких безводных местах (ксероморфные адаптации); об этих приспособлениях речь пойдет в разд. 18.2.3 и 19.3.2.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'