3.4. Отдел Pteridophyta - папоротники, плауны и хвощи
Самые древние папоротникообразные известны нам по ископаемым отпечаткам конца Силурийского периода, их возраст составляет примерно 380 млн. лет. Неизвестно, произошли ли эти растения от мохообразных или независимо от водорослей, но это самые первые из известных нам сосудистых растений. Сосудистые растения - это растения, у которых имеется сосудистая или проводящая ткань, т. е. проводящие ткани ксилемы и флоэмы. Чтобы подчеркнуть, насколько крупным достижением по сравнению с простыми проводящими клетками некоторых мохообразных является возникновение проводящей ткани, все сосудистые растения иногда включают в один отдел Tracheophyta с двумя подотделами - папоротникообразных и более прогрессивных семенных растений.
Наличие проводящих тканей - это один из признаков спорофитного поколения, т. е. поколения, которое у мохообразных относительно невелико и зависит от гаметофита. Именно наличие проводящей ткани не у гаметофитного, а у спорофитного поколения является единственной причиной, по которой у всех сосудистых растений доминирует спорофит.
Для нас здесь важны два свойства проводящей ткани. Во-первых, она образует транспортную систему, которая разносит питательные вещества и воду по всем клеткам тела, что позволяет растениям достигать больших размеров и сложной организации. Во-вторых, тело растения получает внутреннюю опору, так как ксилема не только служит проводящей тканью, но и содержит лигнифицированные клетки, которые обладают большой прочностью и жесткостью. У некоторых вымерших папоротникообразных за счет вторичного роста интенсивно развивалась ксилема, образуя древесину - основную опорную ткань современных деревьев и кустарников. У сосудистых растений развита и другая лигнифицированная ткань - склеренхима, которая дополнительно увеличивает механическую прочность ксилемы (разд. 8.2.1). Проводящие ткани папоротникообразных примитивнее по своему строению, чем проводящие ткани цветковых растений. Так, ксилема папоротникообразных образована не сосудами, а трахеидами, а флоэма - не ситовидными трубками, а ситовидными клетками (разд. 8.2.2).
У самых первых сосудистых растений - псилофитов (эта группа сейчас почти целиком вымерла) - не было корней, которые появились только позднее у других папоротникообразных. Корни проникают глубоко в почву, что облегчает добывание воды, которая разносится по ксилеме ко всем остальным частям растения. В процессе эволюции возникли три явно отличающиеся друг от друга группы папоротникообразных - плауны, хвощи и папоротники; все они дожили до наших дней.
Как только тело растения получило возможность возвышаться над землей, сразу же возникла конкуренция за свет и проявилась тенденция к развитию все более высоких форм. Последовавший за силурийским девонский период отмечен появлением "древовидных" папоротников, одревесневшие стволы которых имели толщину до 2 м и достигали 30 м в высоту. К следующему, каменноугольному периоду повсюду были распространены громадные заболоченные леса из гигантских плаунов и хвощей; из этих "деревьев" в конце концов возникли современные залежи каменного угля. В этих лесах достигли расцвета насекомые и земноводные. Повсюду встречались и папоротники, и древовидные папоротники (у которых не было древесины). Папоротникообразные доминировали среди растительности около 70 млн. лет, начиная с девонского периода и вплоть до пермского периода, когда их вытеснили сначала голосеменные, а затем и цветковые растения (см. геохронологическую шкалу в приложении 5).
Несмотря на большой прогресс в приспособлении спорофитного поколения в сухопутной среде, адаптация практически не затронула гаметофита. Гаметофит у папоротникообразных еще меньше и еще менее стойкий к обезвоживанию, чем у мохообразных; он называется заростком и отмирает тотчас же, как только из него образуется новый спорофит. На заростках развиваются сперматозоиды, которые, плавая в каплях воды, достигают женских гамет.
Разноспоровость
У некоторых папоротникообразных гаметофит защищен и остается внутри споры предшествующего спорофитного поколения. В этом случае образуются споры двух разных типов, поэтому растения с такими особенностями называют разноспоровыми. Растения, у которых все споры одинаковые, как у мохообразных, называют равноспоровыми.
У разноспоровых растений образуются крупные споры, которые называются мегаспорами, и мелкие споры, называемые микроспорами. Структуры, которые участвуют в образовании спор, имеют разные названия, что отражено в табл. 3.6 и на рис. 3.26.
|
Стробил или спороносная шишка - собранные вместе спорофиллы.
Спорофилл лист, на котором образуются спорангии (от греч. Phyllon - лист). Мегаспорофилл - лист, несущий мегаспорангии. Микроспорофилл - лист, несущий микроспорангии. Спорангий - структура, в которой у растений образуются споры; спорангии участвуют в бесполом размножении. Мегаспорангии спорангий, в котором образуются мегаспоры. Микроспорангий спорангий, в котором образуются микроспоры. Мегаспора - сравнительно крупная спора, которая прорастая образует женский гаметофит. Микроспора - относительно мелкая спора, которая прорастая образует мужской гаметофит. Равноспоровое (гомоспоровое) растение-растение, образующее споры только одного типа, например Pellia, Funaria, Dryopteris. Разноспоровое (гетероспоровое) растение-растение, образующее споры двух разных типов, т. е. мегаспоры и микроспоры, например Selaginella и все тайнобрачные растения. |
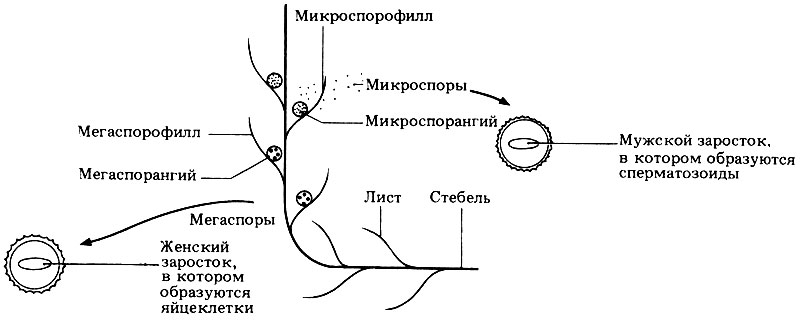
Рис. 3.26. Схема, иллюстрирующая принцип разноспоровости
Из мегаспор вырастают женские гаметофиты (женские заростки), на которых развиваются архегонии, а микроспоры дают начало мужским гаметофитам (мужским заросткам), на которых развиваются антеридии. Сперматозоиды, образующиеся в антеридиях, затем мигрируют к женским заросткам. И мужские, и женские заростки остаются спрятанными внутри спор. Микроспоры очень мелкие, образуется их очень много, и они легко разносятся ветром; вместе с микроспорами рассеиваются и мужские заростки, находящиеся в них. Появление разноспоровости - очень важный этап эволюции семенных растений, о чем мы поговорим ниже.
3.4.1. Систематика и основные признаки Pteridophyta
Систематика и основные признаки современных папоротникообразных представлены в табл. 3.7. Опущен класс Psilopsida, который почти полностью вымер.
3.4.2. Класс Pteropsida - папоротники
Основные признаки Pteropsida перечислены в табл. 3.7. Папоротники обычно встречаются только во влажных тенистых местах. Немногие папоротники могут расти на открытом месте; таким исключением является самый обычный папоротник-орляк (Pteridium). Папоротники широко распространены в тропических дождевых лесах, где условия (температура, освещение и влажность) для них наиболее подходящие.

Таблица 3.7. Систематика и основные признаки Pteridophyta
Мужской папоротник (Dryopteris filix-mas), пожалуй, наиболее распространен в Великобритании; он встречается по всей стране в сырых лесах, лесополосах и других тенистых местах. Вайи (листья) спорофита достигают высоты до 1 м и более и растут от толстого горизонтального стебля или корневища. На корневище находятся придаточные корни. От основного стебля могут отламываться отдельные ветви и давать начало новым растениям, это - одна из форм вегетативного размножения. У основания слоевище покрыто сухими бурыми чешуйками, которые защищают молодые листья от заморозков и от засухи. Молодые листья плотно закручены в характерные для папоротников "завитки". Выше по слоевищу размеры чешуек постепенно уменьшаются, и они все дальше отходят друг от друга. Главная ось слоевища называется главным черешком, а листочки, отходящие от нее в обе стороны, называются листочками перистого листа. Небольшие округлые выступы на листочках называются вторичными листочками. С внешним видом и основными признаками спорофита Dryopteris filix-mas можно познакомиться на рис. 3.27.
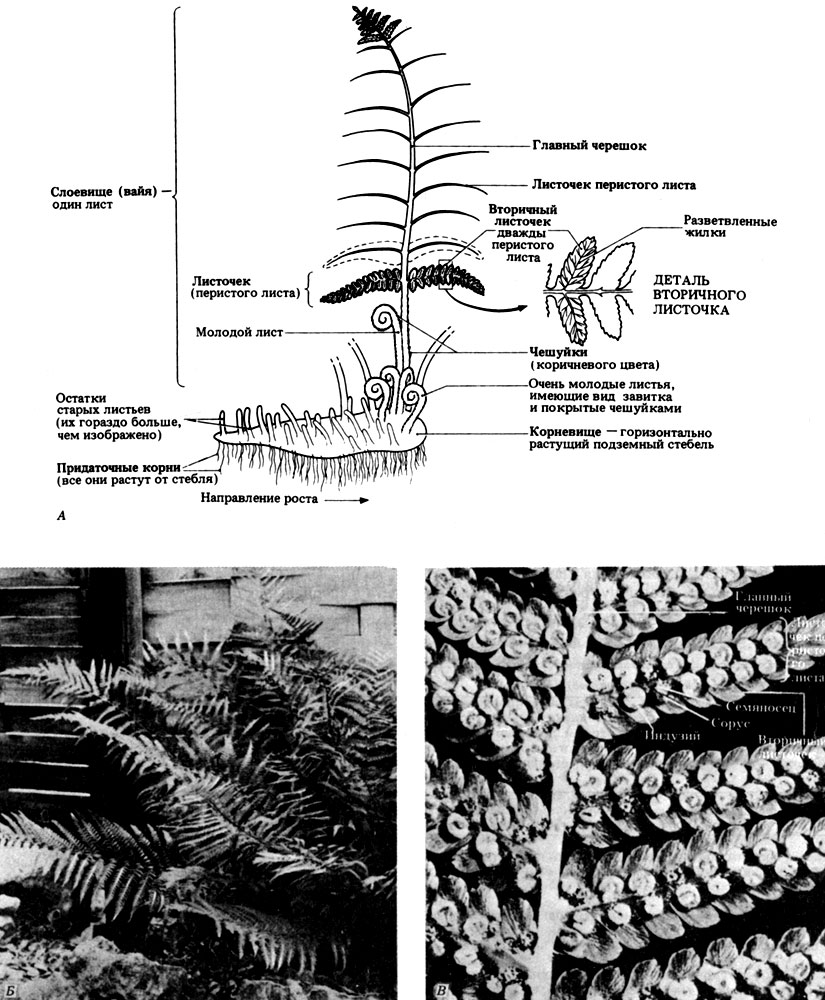
Рис. 3.27. Внешний вид и основные признаки спорофитного поколения мужского папоротника, или щитовника (Dryopteris filix-mas). А. Схема и детали строения одной пары листочков; все другие имеют такое же строение. Б. Листья папоротника. В. Нижняя поверхность листа с видимыми на ней сорусами (некоторые сорусы покрыты индузием)
Жизненный цикл
Жизненный цикл Dryopteris представлен в виде схемы на рис. 3.28.
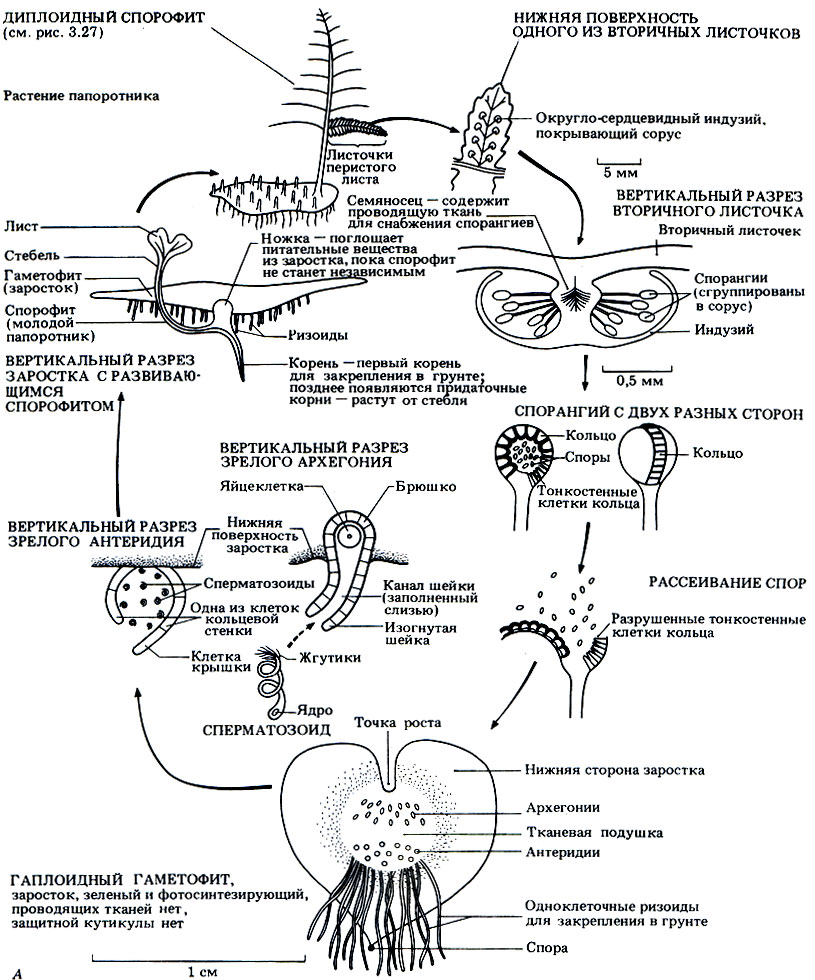
Рис. 3.28. А. Жизненный цикл Dryopteris filix-mas
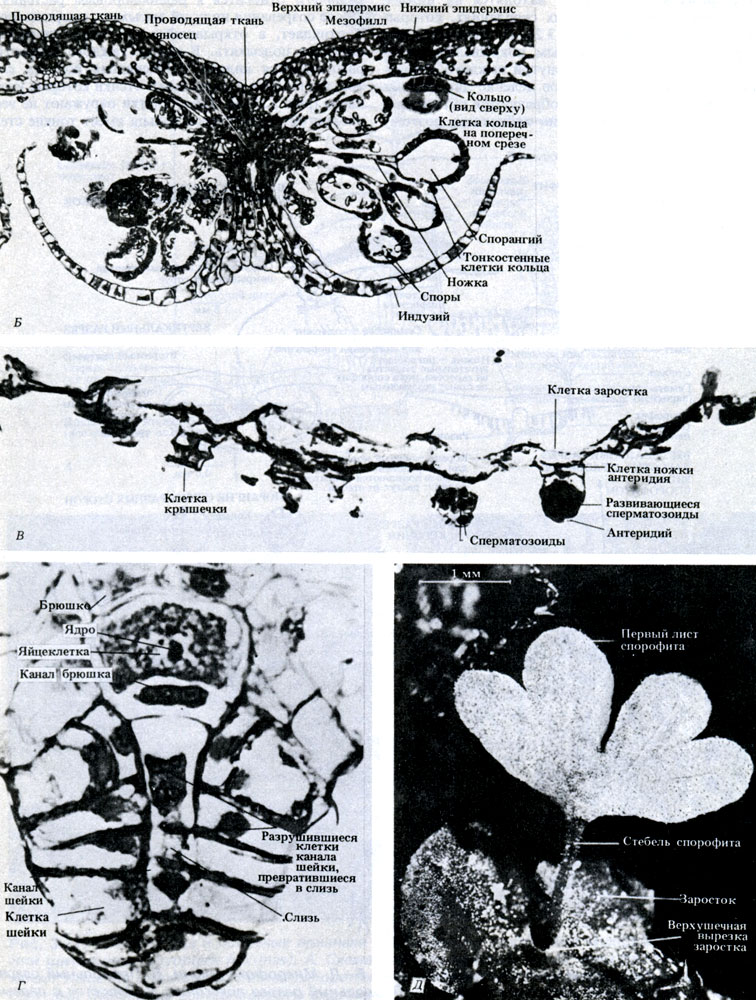
Рис. 3.28. Б-Д. Микрофотографии. Б. Продольный разрез соруса. В. Продольный разрез антеридиев. Г. Продольный разрез архегония. Д. Заросток с первым появляющимся листом
Бесполое размножение. Споры образуются в конце лета в специальных структурах, которые называются спорангиями. Спорангии находятся на нижней поверхности листа в особых скоплениях, которые называются сорусами (рис. 3.28, А). Каждый сорус закрыт округло-сердцевидным покрывалом, которое называется индузием. Внутри каждого спорангия происходит мейотическое деление диплоидных материнских клеток спор и образуются гаплоидные споры. Все споры совершенно одинаковы, поэтому Dryopteris относится к равноспоровым растениям. После созревания индузий высыхает, сморщивается и отпадает, а открывшийся при этом спорангий начинает подсыхать. В стенке каждого спорангия располагается кольцо - гребневидная полоска клеток, внутренние и радиальные стенки которых утолщены (рис. 3.28, Б). Такие клетки окружают не весь спорангий; часть клеток кольца имеет тонкие стенки. Несколько тонкостенных клеток образуют особый участок, называемый стомиумом. По мере высыхания клеток кольца их тонкие наружные стенки начинают втягиваться внутрь сморщенной цитоплазмы. Возникающее при этом напряжение заставляет лопаться всю полоску тонкостенных клеток, и при этом кольцо закручивается назад. Когда клетки лопаются, споры "выстреливают" из спорангия, как из катапульты. В конце концов цитоплазма тоже выталкивается из клеток кольца, в результате чего в нем резко снимается напряжение, и оно снова возвращается в исходное положение, выбрасывая наружу последние остатки спор.
Прорастание. Споры остаются в состоянии покоя недолго, и если условия позволяют, то они прорастают и дают начало новому гаметофитному поколению. Гаметофит представляет собой тонкую сердцевидную пластинку диаметром около 1 см (рис. 3.28, А). Пластинка зеленая, способна к фотосинтезу и прикрепляется к почве одноклеточными ризоидами. У такого очень нежного заростка нет кутикулы, он быстро высыхает и поэтому может жить только там, где достаточно влаги.
Половое размножение. На нижней поверхности гаметофита (заростка) образуются простые антеридии и архегонии (рис. 3.28, А). Эти репродуктивные органы защищают находящиеся в них гаметы. Гаметы возникают путем митоза из материнских клеток гамет; при этом, как и у мохообразных, в антеридиях образуются сперматозоиды, а в архегониях - по одной яйцеклетке. У каждого сперматозоида имеется пучок жгутиков. Если влаги хватает, то из антеридиев выходят созревшие сперматозоиды, которые подплывают к архегониям. Движение сперматозоидов обусловлено ответной реакцией хемотаксиса на яблочную кислоту (2-гидроксибутан-дикарбоновую кислоту), выделяемую клетками шейки архегония. Оплодотворение обычно перекрестное, потому что антеридии созревают раньше архегониев. После оплодотворения образуется диплоидная зигота. Запомните, что оплодотворение, как и у мохообразных, все еще зависит от наличия воды.
Развитие зиготы. Из зиготы развивается спорофит. У молодого зародыша образуется ножка, через которую он поглощает питательные вещества гаметофита, пока эту функцию не возьмут на себя собственные корни и листья спорофита. Гаметофит вскоре увядает и отмирает.
Жизненный цикл папоротника схематически представлен на рис. 3.29.
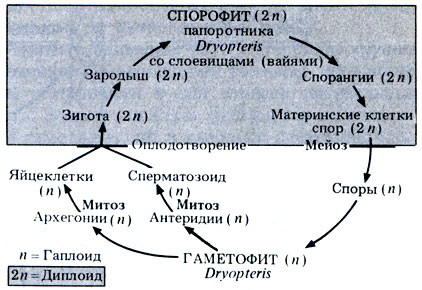
Рис. 3.29. Схема жизненного цикла Dryopteris filix-mas
3.7. В чем выражается более совершенная, чем у печеночников и мхов, адаптация папоротников к жизни на суше?
3.8. Какие поколения папоротникообразных могут полностью обеспечивать себя питательными веществами?
Выберите правильные ответы:
а) взрослые печеночники и гаметофиты мхов;
б) взрослые печеночники и спорофиты мхов;
в) взрослые гаметофиты папоротников;
г) взрослые спорофиты папоротников.
3.9. В чем выражается недостаточная адаптация мхов, печеночников и папоротников к жизни на суше?
3.10. Как распространяются папоротники?
3.11. а) Чем защищена зигота печеночников (или мхов) и папоротников? б) Как она снабжается питательными веществами?
3.4.3. Класс Lycopsida - плауны
Основные признаки Lycopsida перечислены в табл. 3.7. Обратите внимание на то, что, несмотря на некоторое внешнее сходство с мхами, эти растения принадлежат к папоротникообразным и более высоко организованы, чем настоящие мхи, которые относятся к мохообразным. Когда-то Lycopsida были распространены гораздо шире, чем в наши дни; среди них было много древовидных, о чем говорилось на с. 76. По своей приспособленности к жизни на суше плауны занимают промежуточное положение между папоротниками и семенными растениями.
Представители рода Selaginella (плаунки) распространены главным образом в тропиках, а в Великобритании встречается всего один вид - S. selaginoides. Это растение довольно обычно в горных областях на северо-западе Великобритании, где его можно найти в благоприятных для него влажных условиях, на мокрых скалах, пастбищах и вблизи воды. Для S. selaginoides характерен стелющийся стебель, который обычно плоско лежит на земле, и от него отходят короткие прямостоячие ветви. Внешний вид самой обычной селагинеллы, часто встречающейся в теплицах, - S. Kraussiana - показан на рис. 3.30. У этого растения четыре ряда небольших листьев, которые расположены супротивно парами, при этом каждая пара состоит из большого (нижнего) и малого (верхнего) листьев. У основания каждого листа имеется язычок (лигула) - небольшой пленчатый вырост. Вниз от стебля отходят похожие на корни образования, которые называются ризофорами. Ризофоры ветвятся и дают придаточные корни.
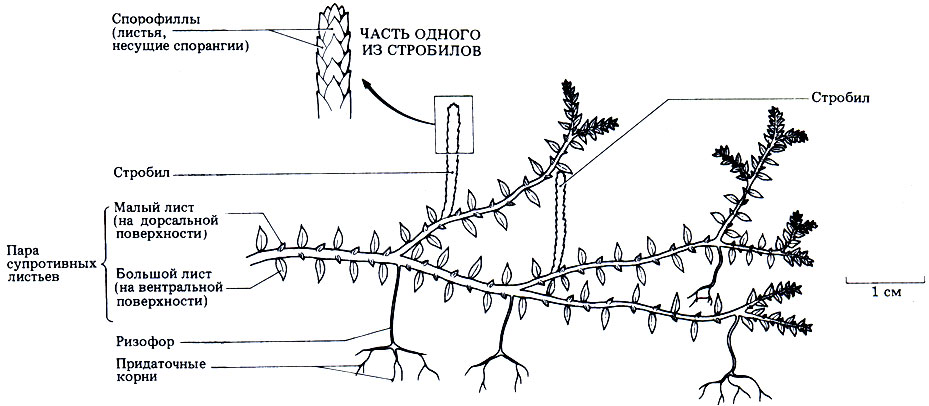
Рис. 3.30. Внешний вид и характерные признаки спорофитного поколения Selaginella kraussiana - растения, обычно встречающегося в теплицах. У Selaginella selaginoides - единственного вида, растущего в Великобритании, - все листья одинакового размера. Кроме того, у него нет ризофоров, а есть только придаточные корни
При размножении образуются вертикальные ветви, которые называются стробилами или шишками. Стробилы состоят из четырех вертикальных рядов листьев одинакового размера, на дорсальной поверхности которых образуются спорангии; поэтому их называют спорофиллами.
Жизненный цикл
Жизненный цикл Selaginella изображен схематически на рис. 3.31. Если вам не известно значение терминов, которые мы будем использовать дальше, обратитесь к табл. 3.6.
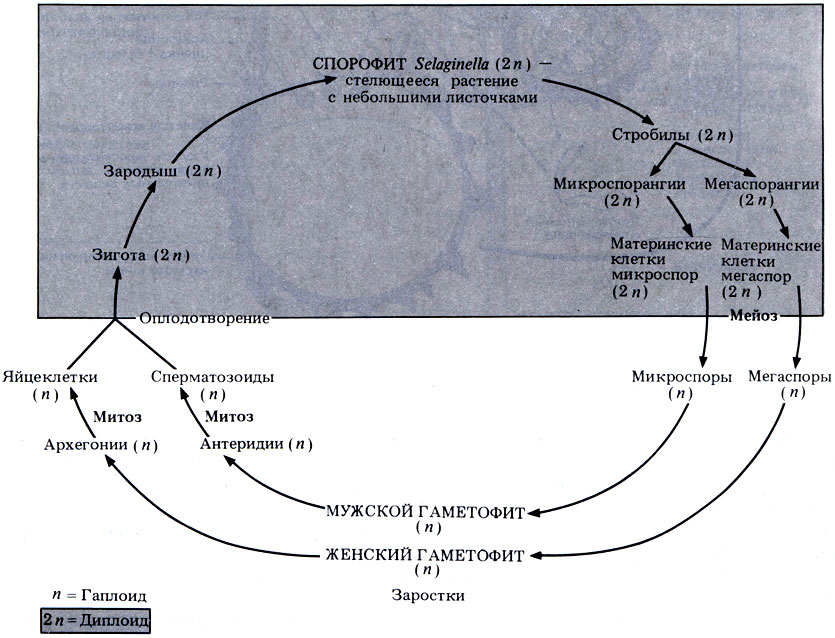
Рис. 3.31. Схема жизненного цикла Selaginella
Бесполое размножение. Как сказано выше, Selaginella образует стробилы, или шишки. Нижние листья представляют собой мегаспорофиллы, на которых образуются мегаспорангии, верхние листья - микроспорофиллы, на которых образуются микроспорангии (рис. 3.32). В каждом мегаспорангии образуются по четыре мегаспоры, а в каждом микроспорангии - множество микроспор; и в том и в другом случае происходит мейотическое деление материнских клеток спор. Поскольку образуются споры двух разных типов, Selaginella относят к разноспоровым растениям.
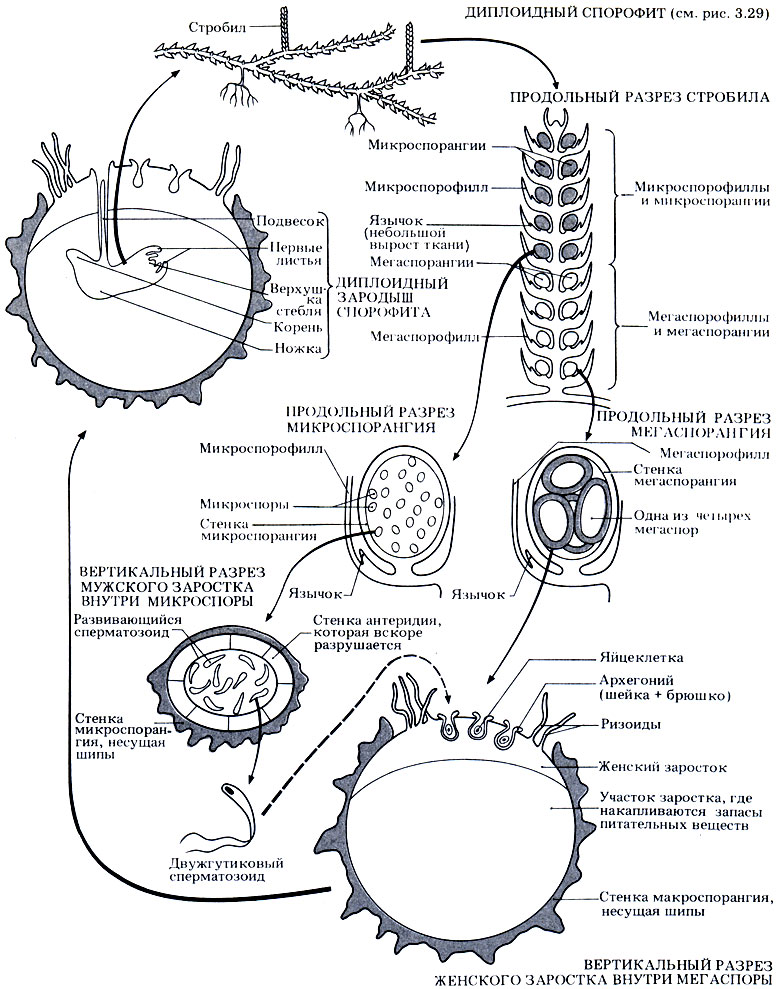
Рис. 3.32. Жизненный цикл Selaginella
Развитие спор и половое размножение. микроспоры дают начало мужским гаметофитам. В процессе развития микроспоры высвобождаются и рассеиваются или же высыпаются вниз на мегаспорофиллы. Содержимое микроспоры превращается в мужской заросток, состоящий из одной вегетативной клетки и одиночного антеридия, внутри которого после митоза образуются сперматозоиды со жгутиками. Заросток - редуцированное гаметофитное поколение - не способен к фотосинтезу и целиком зависит от запаса питательных веществ в микроспоре. Можно проследить, что эти вещества образовались еще у предыдущего спорофитного поколения.
Из мегаспор развиваются женские гаметофиты. Снова развитие начинается задолго до рассеивания спор, причем содержимое мегаспоры превращается в женский заросток - редуцированный гаметофит. После раскрывания споры заросток выходит наружу. Он образует ризоиды, частично зеленеет, и в нем начинаются процессы фотосинтеза. Однако, как и у мужских гаметофитов, основная часть питательных веществ поступает из запасов споры, которые образовались еще у предшествующего спорофитного поколения. На поверхности женского заростка появляются архегонии, внутри которых находится по одной яйцеклетке, образовавшейся путем митоза.
Обратите внимание на то, что зрелые гаметофиты плаунов не отделены от спорофита и этим отличаются от всех уже рассмотренных нами сухопутных растений. Это очень важное эволюционное достижение - одно из приспособлений к жизни на суше, которое привело к тому, что легко уязвимый гаметофит стал хотя бы отчасти защищен спорой. Гаметофитное поколение не способно жить самостоятельно, оно питается за счет веществ, которые запасло в споре предшествующее спорофитное поколение.
Вместе с преимуществами проявились и сопутствующие им недостатки, связанные с тем, что сперматозоиды должны путешествовать от мужского заростка, находящегося внутри микроспоры, к женскому заростку, лежащему внутри мегаспоры. Самооплодотворение гаметофита становится совсем невозможным, а споры (а следовательно, и гаметофиты) при рассеивании могут быть отнесены далеко друг от друга.
Оплодотворение. Стенки микроспор разрушаются, и сперматозоиды выходят наружу. Для того чтобы это произошло, нужна вода, поскольку только тогда сперматозоиды смогут подплыть к архегониям на женском заростке. Заросток все еще находится в мегаспоре, а она либо по-прежнему прикреплена к родительскому спорофиту, либо же отделяется от него. Сперматозоиды проникают в шейку архегония, один из них сливается с яйцеклеткой, и образуется диплоидная зигота.
Развитие зиготы. Зигота развивается в зародыш спорофита. Верхняя часть зародыша превращается в удлиненную структуру, называемую подвеском, которая проталкивает зародыш к запасу питательных веществ гаметофита и мегаспоры. У зародыша образуются корень, стебель и листья. До тех пор, пока он не станет самостоятельным фотосинтезирующим растением, он питается через ножку.
Обратите внимание на то, что более крупные по сравнению с микроспорой размеры мегаспоры связаны с наличием в ней запасов питательных веществ. Этими запасами снабжается не только женский гаметофит, но и зародыш следующего за ним спорофитного поколения. Таким образом, питательные вещества, запасенные одним спорофитным поколением, используются на ранних этапах развития следующего спорофитного поколения. Жизненный цикл Selaginella показан на рис. 3.32.
3.4.4. Класс Sphenopsida - клинолистовидные, или членистые (хвощи)
Основные признаки Sphenopsida представлены в табл. 3.7. Единственный дошедший до нас род Equisetum включает в себя около 25 видов*, распространенных по всему земному шару (кроме Австралии)**. Многие из них обитают во влажных и сырых местах, вблизи прудов и на болотах. Однако обыкновенный, или полевой, хвощ (Equisetum arvense) обычен по всей Великобритании и встречается даже в более сухих местах: на полях, по обочинам дорог, на пустошах и в садах.
* (32 вида. - Прим. перев.)
** (И Новой Зеландии. - Прим. перев.)
У спорофитов имеются горизонтальные подземные стебли (корневища), а высота надземных побегов обычно не превышает 1 м. От узлов отходят мутовки мелких заостренных листьев, похожих на чешуйки. Воздушные побеги бывают двух типов - "стерильные" вегетативные побеги и "плодущие" побеги, несущие спороносные шишки (стробилы). Стерильные побеги зеленые; от узлов у них отходят не только мутовки чешуевидных листьев, но и мутовки ветвей. Плодущие побеги у большинства видов бесцветные или бледнобурые, не ветвятся, и на верхушке у них сидит по одному спороносному стробилу; у некоторых видов эти побеги зеленые и ветвятся. Междоузлия (участки между узлами) всех составных частей растения в середине полые, а снаружи покрыты рядом продольных низбегающих бороздок. Внешний вид и некоторые признаки Е. arvense изображены на рис. 3.33.
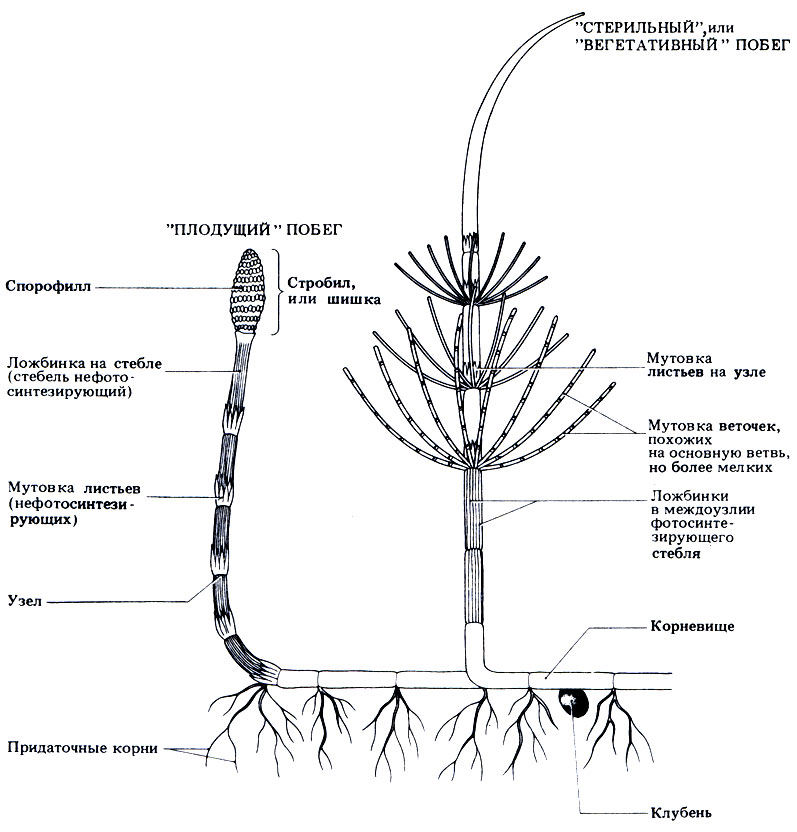
Рис. 3.33. Внешний вид и характерные особенности спорофитного поколения Equisetum arvense. 'Плодущие' (спорангиеносные) побеги высотой 10-25 см красно-коричневого цвета появляются в марте. Спорофилл представляет собой плоский диск на короткой ножке, на нижней стороне которого располагается 5-10 спорангиев. Стерильные вегетативные побеги достигают в высоту 20-80 см. На каждом узле побега располагается мутовка мелких, похожих на чешуйки зеленых листьев (микрофиллов). Корневище представляет собой горизонтальный подземный стебель с хорошо выраженными узлами и междоузлиями; на каждом узле корневища развиваются хорошо разветвленные придаточные корни. Клубень - это короткая округлая ветвь длиной в одно междоузлие; предназначен для зимовки и вегетативного размножения
Советуем вам увидеть огромный каталог проституток в этом популярном сайте для мужчин prostitutkitverigirls.com, если вы интересуетесь возможностью встретиться с любимыми индивидуалками региона.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'