Глава 9
9.1. Фотоавтотрофное питание - это процесс, в котором световая энергия Солнца служит источником энергии для синтеза органических соединений из неорганических веществ с использованием СО2 как источника углерода. Хемогетеротрофное питание - это процесс, в котором органические соединения синтезируются из уже существующих органических источников углерода за счет энергии химических реакций.
9.2. а) Автотрофный тип питания; б) Гетеротрофный тип питания.
9.3. 75·1012 кг углерода в год [(40·1012)+(35·1012)].
9.4. Энергия солнца доступна; сырье - вода - имеется в большом количестве; продуктом горения также является вода, которая не токсична и не загрязняет окружающую среду (атомная энергия небезопасна и является источником загрязнения среды).
9.5. Общая форма и положение
Большое отношение площади поверхности к объему для максимального улавливания света и эффективного газообмена. Листовая пластинка, как правило, располагается под прямым углом к падающему свету, особенно у двудольных растений.
Устьица
Отверстия в листьях, обеспечивающие газообмен. При фотосинтезе поглощается двуокись углерода и как побочный продукт выделяется кислород. У двудольных растений устьица расположены в основном на нижней, теневой, стороне листа, таким образом обеспечивается минимальная потеря воды в результате транспирации.
Замыкающие клетки
Регулируют открывание устьиц (обеспечивают открывание устьиц только на свету, когда идет фотосинтез).
Мезофилл
Содержит специальные органеллы для фотосинтеза-хлоропласта, в которых находится хлорофилл. У двудольных растений палисадные клетки мезофилла, содержащие наибольшее число хлоропластов, расположены непосредственно под верхней поверхностью листа, что обеспечивает максимальное улавливание света. Палисадные клетки вытянуты, что увеличивает вероятность поглощения ими света. Хлоропласты расположены по периферии клеток, что облегчает газообмен с межклеточным пространством. Хлоропласты могут обладать фототаксисом (т. е. перемещаться внутри клетки по направлению к свету). У двудольных растений губчатый мезофилл содержит крупные межклеточные пространства, что обеспечивает эффективность газообмена. (Листья однодольных растений также содержат крупные межклеточные пространства.)
Проводящая система
Снабжает водой, участвующей в процессе фотосинтеза, а также минеральными солями. Транспортирует продукты фотосинтеза. Колленхима и склеренхима служат опорой растению.
9.6. Хлорофилл а поглощает красный свет примерно вдвое интенсивнее, чем хлорофилл b, а его максимум поглощения приходится на несколько более длинные волны (волны с меньшей энергией). Поглощение в синем свете идет слабее и смещено к несколько более коротким волнам (волнам с большей энергией). Обратите внимание, что эти различия вызваны очень незначительными изменениями в химической структуре.
9.7. Хлорофилл а имеет более низкую энергию возбуждения. Таким образом хлорофилл b может передать возбужденные электроны (эквивалентно энергии) на хлорофилл я, несмотря на то что при переносе некоторая часть энергии теряется в виде тепла.
9.8. Темно-синий цвет красителя по мере его восстановления должен исчезнуть, оставив зеленый цвет хлоропластов.
9.9. ДХФИФ должен остаться синим в контрольных пробирках 2 и 3. На примере пробирки 2 показано, что одним светом нельзя вызвать изменения цвета и что для осуществления реакции Хилла необходимы хлоропласты. На примере пробирки 3 видно, что для протекания реакции Хилла свет необходим так же, как и хлоропласты.
9.10. Двумя органеллами, наиболее близкими к хлоропластам по размерам, являются ядра (немного больше) и митохондрии (немного меньше). Для того чтобы выделить чистые хлоропласты, необходимо более точное дифференциальное центрифугирование или центрифугирование в градиенте плотности.
9.11. Косвенные данные позволяют предположить, что ядра и митохондрии не участвуют в восстановлении ДХФИФ, так как для этого необходим свет, а эти органеллы не содержат ни хлорофилла, ни любого другого видимого пигмента.
9.12. Для того чтобы уменьшить активность фермента. Во время гомогенизации ферменты, разрушающие химические соединения клетки, могли высвободиться из других частей клетки, например из лизосом или вакуолей.
9.13. Клеточные реакции проходят эффективно только при определенных рН; любое существенное изменение рН, вызванное, например, высвобождением кислот из других частей клетки, может повлиять на активность хлоропластов.
9.14. а) Вода; б) ДХФИФ.
9.15. Только с нециклическим фотофосфорилированием: 1) выделился кислород; 2) электроны были захвачены ДХФИФ, поэтому они не могли вернуться в ФС I.
9.16. а) У хлоропластов отсутствуют оболочка (ограничивающая мембрана) и строма. Осталась только внутренняя система мембран.
б) Среда без сахарозы является гипотонической для хлоропластов. Не имея защитной клеточной стенки, разрушенной во время гомогенизации клеток, хлоропласты осмотически поглощают воду, набухают и лопаются. Строма растворяется, остаются только мембраны.
в) Данные изменения желательны, потому что лопнувшие хлоропласты обеспечивают более эффективный доступ ДХФИФ к мембранам, где протекает реакция Хилла.
9.17. Открытие реакции Хилла послужило поворотным пунктом по нескольким причинам:
1) Она показала, что выделение кислорода может происходить без восстановления двуокиси углерода, доказывая тем самым, что световые и темновые реакции и реакции расщепления воды разобщены.
2) Она показала, что хлоропласты могут осуществлять светозависимое восстановление акцепторов электронов.
3) Она предоставила биохимические данные о том, что световая реакция фотосинтеза полностью сосредоточена в хлоропласте.
9.18. Изотоп с непродолжительным периодом полураспада (например, 11С, 20,5 мин) быстро распадается, так что его уже нельзя обнаружить; следовательно, его пригодность для биологических экспериментов, которые зачастую требуют для своего завершения нескольких часов или дней, существенно ограничивается.
9.19. Биохимические реакции, протекающие в процессе фотосинтеза у Chlorella и у высших растений, сходны; по этой и по ряду приведенных ниже причин при изучении фотосинтеза обычно используется Chlorella:
1) Культура Chlorella фактически является культурой хлоропластов, так как большая часть объема каждой клетки занята отдельным хлоропластом.
2) В культуре удается, получить более однородный рост.
3) Клетки хлореллы очень быстро подвергаются действию радиоактивной двуокиси углерода и столь же быстро погибают, поэтому техника ухода за культурой проста.
9.20. Для максимального освещения водорослей.
9.21.
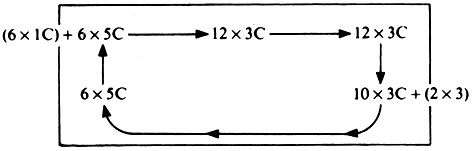
Схема
Этой схемой подчеркивается циклический перенос углерода; сложность цикла Кальвина обусловлена главным образом трудностью превращения 10·3С в 6·5С.
9.22. Наличие двуокиси углерода, воды, света и хлорофилла.
9.23. а) На участке А лимитирующим фактором является интенсивность света.
б) Б: фактор, отличный от интенсивности света, становится лимитирующим фактором. На участке Б лимитирующими являются как интенсивность света, так и другой (ие) фактор (ы). В: интенсивность света больше не является лимитирующим фактором.
в) Г: "точка насыщения" для интенсивности света в этих условиях, т. е. точка, за которой усиление интенсивности света не вызывает дальнейшего увеличения скорости фотосинтеза.
г) Д: максимальная скорость фотосинтеза, достижимая в условиях эксперимента.
9.24. X, Y и Z - точки, в которых свет перестает быть главным лимитирующим фактором в четырех экспериментах. Выше этих точек существует линейная зависимость между интенсивностью света и скоростью фотосинтеза.
9.25. Ферменты начинают денатурировать.
9.26. Подобные условия возникают а) в затененном сообществе, например в лесу, на рассвете и в сумерках в теплом климате; б) обычно лимитирующим фактором служит содержание СО2, но и такие условия могут возникнуть особенно в сомкнутых насаждениях растений, например в посевах в солнечных и теплых условиях; в) в ясный зимний день.
9.27. Растения в темноте продолжают расходовать сахара, например для дыхания. В темноте фотосинтез прекращается, и, после того как израсходуются все сахара, в сахара превращается запасенный крахмал, который расщепляется, в частности, до сахарозы, транспортируемой от листьев к другим частям растений.
9.28. Можно оспорить предположение, что бумага или фольга препятствуют прохождению фотосинтеза, ограничивая проникновение СО2 к закрытым частям листа. Это предположение можно опровергнуть, оставив воздушную щель между бумагой и листом, как это показано ниже на рис. 9.28 (отв.).
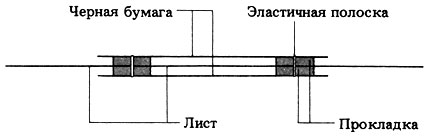
Рис. 9.28. (отв.). Срез листа, завернутого в черную бумагу
9.29. Его необходимо поместить в такую же колбу, но воду заменить раствором гидроксида калия. Непромокаемая вата будет защищать стебель, на котором находится лист. (Поверхность самого стебля можно обработать известковой водой, чтобы воспрепятствовать возможному повреждению, которое может повлиять на фотосинтез.)
9.30. Можно измерять скорость поглощения СО2, скорость выделения кислорода и скорость образования углеводов. Можно также измерять скорость прироста сухой массы листьев. Такое измерение особенно удобно проводить на культурных растениях во время сезона вегетации, когда можно отобрать сравнительно крупные образцы. Опыт по измерению скорости поглощения СО2 описан в разд. 9.7.
9.31. а) Скорость образования газа прямо пропорциональна I вплоть до значения I, равного х-единицам. В этой точке начинается насыщение светом и завершается в точке у (значения х и у зависят от условий опыта). Следовательно, скорость образования газа лимитировалась не светом, а каким-то другим фактором. б) Лабораторию затенили для того, чтобы предотвратить попадание света снаружи, который мог бы стимулировать дополнительный фотосинтез. Температуру поддерживают постоянной, поскольку ее изменения также влияют на скорость фотосинтеза.
9.32. а) Может изменяться температура, поскольку лампа нагревает воду (этого следует избегать путем использования водяной бани).
б) Во время эксперимента может изменяться концентрация СО2 в воде, особенно если ранее был добавлен К2СО3.
в) Любой случайно попавший в лабораторию свет будет влиять на фотосинтез.
9.33. По мере того как пузырьки кислорода поднимаются к поверхности воды, часть растворенного в воде азота переходит из раствора в эти пузырьки, а часть кислорода пузырьков растворяется в воде. Этот обмен происходит вследствие различий давления (концентрации) кислорода и азота в пузырьках и в воде; со временем концентрации этих веществ стремятся прийти в равновесие. В собранном газе в следовых количествах также будут присутствовать водяные пары и СО2. Собранный газ будет стремиться прийти в равновесное состояние с атмосферным воздухом посредством диффузии газов через воду.
9.34. Необходимо собрать весь кислород, выделенный в процессе фотосинтеза за время опыта. Если вода не будет аэрирована, то часть выделяющегося при фотосинтезе кислорода будет растворяться в воде и, следовательно, будет зарегистрировано меньшее количество кислорода.
9.35. Образец записи результатов приведен в следующей таблице:

Таблица
Контрольные пробирки В и Г необходимы для того, чтобы подтвердить, что любые изменения в пробирках А и Б происходят только при наличии в них листьев. В пробирке А среда становится более кислой из-за выделяющейся во время дыхания СО2. В отсутствие света фотосинтез не идет. В пробирке Б среда становится менее кислой, что указывает на расходование СО2. СО2, выделяющаяся во время дыхания, использовалась при фотосинтезе совместно с СО2, которая уже находилась в окружающем лист воздухе, и была растворена в растворе индикатора. Скорость фотосинтеза была выше скорости дыхания.
9.36. Компенсационная точка для СО2. В этой точке скорость фотосинтеза равна скорости дыхания.
9.37. При увеличении концентрации СО2 до 0,1% скорость фотосинтеза возрастает. С возрастанием концентрации СО2 более успешно конкурирует с кислородом за активный центр на РиБФ-карбоксилазе, следовательно, увеличивается скорость фиксации СО2, т. е. скорость фотосинтеза. Увеличение концентрации кислорода тормозит фотосинтез по противоположной причине. Кислород стремится вытеснить двуокись углерода и стимулирует фотодыхание, при котором выделяется СО2.
9.38. Высокая концентрация кислорода и низкая концентрация СО2 (максимальная скорость достигается при 100%-ной концентрации кислорода). Для достижения большой скорости необходима также высокая интенсивность света, поскольку процесс является светозависимым.
9.39. Хлоропласты мезофилла участвуют в световых реакциях, хлоропласта обкладки проводящих пучков участвуют в темновых реакциях.
9.40. Образование кислорода связано с гранами (основное местоположение ФС II); кислород конкурирует с СО2 за РиБФ-карбоксилазу и стимулирует фото дыхание. Кроме того, граны занимают большой объем хлоропласта, а если их нет, то больший объем занимают строма и соответственно РиБФ-карбоксилаза, поэтому остается место для откладывающегося крахмала.
9.41. Насос для перекачки двуокиси углерода. Малатный шунт, действуя как насос для перекачки двуокиси углерода, увеличивает ее концентрацию в клетках обкладки проводящих пучков, увеличивая таким образом эффективность работы РиБФ-карбоксилазы. Насос для перекачки водорода. Малат переносит водород от НАДФ·Н2, содержащегося в мезофилле, к НАДФ в клетках обкладки проводящих пучков, где восстанавливается НАДФ·Н2. Преимущество подобного переноса состоит в том, что НАДФ·Н2 образуется в результате эффективной световой реакции в хлоропластах мезофилла (присутствует ФС II) и затем может быть использован в качестве восстановительного потенциала в цикле Кальвина в хлоропластах клеток обкладки проводящих пучков, в которых собственный синтез НАДФ·Н2 ограничен.
9.42. а) Понижение концентрации кислорода стимулирует С3-фотосинтез, так как при этом уменьшается фото дыхание.
б) Понижение концентрации кислорода не влияет на С4-фотосинтез, потому что фото дыхание уже ингибировано.
9.43. Симбиотические бактерии, находящиеся в корневых клубеньках растений из семейства бобовых, связывают азот. Это приводит к усилению роста и соответственно к увеличению потребности в других минеральных веществах, особенно в калии и фосфоре (иногда посевы бобовых перепахивают, удерживая таким образом минеральные вещества в почве).
9.44. Хемогетеротрофному. Далее их можно классифицировать как сапрофитные.
9.45. Любые, в которых количество кислорода недостаточно для того, чтобы разложить всю накопленную массу органического вещества, например на болотах, в донных осадках, таких, как ил, в арктической тундре, в глубоких горизонтах почвы и в полузатопленных почвах.
9.46. И то и другое увеличивает аэрацию и соответственно содержание кислорода в почве. Это стимулирует разложение и нитрификацию, а также подавляет денитрификацию, поскольку вместо нитратов используется кислород.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'