19.4.8. Позвоночные
У различных позвоночных встречается экскреция всех трех азотистых продуктов, что, как правило, зависит от доступности воды для того или иного вида. Механизмы осморегуляции у позвоночных более эффективны, чем у беспозвоночных, благодаря малой проницаемости наружных покровов и наличию почек. Биологи до сих пор спорят о том, где возникли первые рыбы в морской или пресной воде. Многие биологи считают более вероятным морское происхождение первых рыб и рассматривают почки как более позднее приобретение, необходимое для выживания в гипотонических условиях пресных водоемов. В этих условиях почки служат для удаления избытка воды и задержки солей. Последующее развитие почек зависело от характера окружающей среды и шло по линии все большего усложнения в ряду позвоночных от рыб до млекопитающих. Увеличение сложности строения почек было связано с заселением суши. Благодаря повышенной эффективности механизмов выделения и осморегуляции состав внутренней среды у позвоночных колеблется в более узких пределах, чем у беспозвоночных.
Структурно-функциональной единицей почечной ткани является нефрон. Нефроны - сегментарные структуры, образовавшиеся из мезодермальных нефротомов (разд. 21.8), вступивших в тесный контакт с кровеносными сосудами, отходящими от аорты, и связанных с целомом через ресничную воронку. Нефроны у зародышей рыб имеют наиболее примитивное строение, при котором несколько нефронов открываются в перикардиальную полость и в совокупности образуют структуру, называемую пронефросом (рис. 19.13) или предпочкой. У всех взрослых рыб и амфибий пронефрос утрачивается, а вместо него развивается более компактное образование состоящее из значительно большего числа нефронов и расположенное в брюшном и хвостовом отделах тела. Это мезонефрос, или первичная почка. В мезо- нефросе нефроны утратили связь с целомом и объединены собирательным протоком, ведущим к мочеполовому отверстию. Такое строение идеально подходит для выделения разбавленной мочи, образующейся в основном у обитателей пресных вод.
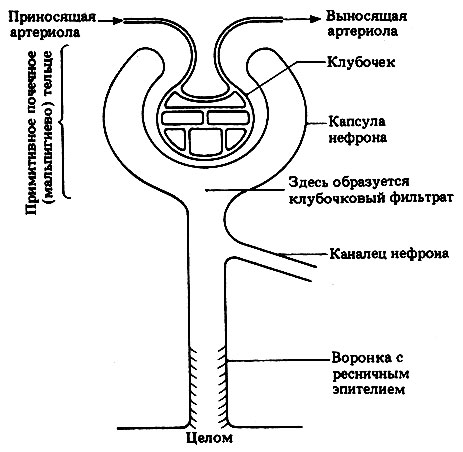
Рис. 19.13. Строение примитивного нефрона. Образующийся в клубочке фильтрат поступает через воронку с ресничным эпителием в целом или через соединительный каналец объединяется с фильтратом из других нефронов
Рептилии, птицы и млекопитающие приспособились к жизни на суше, где вместо проблемы удаления воды, стоящей перед рыбами и амфибиями, встает задача удержания воды в организме. У этих животных орган выделения представляет собой еще более компактную структуру - метанефрос, или вторичную почку, которая состоит из еще большего числа нефронов с еще более длинными канальцами. В канальцах происходит реабсорбция воды и образуется концентрированная моча, которая в конце концов поступает в почечную лоханку, а из нее - в мочевой пузырь. (Более подробно строение и функции почки млекопитающего описаны в разд. 19.5.)
Образование мочи в почке позвоночных основано на принципах ультрафильтрации, избирательной реабсорбции и активной секреции. Моча представляет собой жидкость, содержащую отходы азотистого обмена, воду и те ионы, содержание которых в организме превышает необходимый уровень. Ультрафильтрации подвергаются также и ценные для организма вещества, но они всасываются обратно в кровь. Реабсорбируется 99% растворенных веществ, и на этот процесс расходуется энергия. С энергетической точки зрения такой механизм кажется неэкономным, но он обеспечивает позвоночным большую гибкость при освоении новых мест обитания, так как позволяет выводить чужеродные или "новые" вещества, как только они появляются в организме, и для их удаления не нужно создавать новый секреторный механизм.
У рыб органами выделения и осморегуляции служат жабры и почки. Оба органа проницаемы для воды, азотистых отходов и ионов и обладают большой поверхностью, облегчающей обмен. Почки в отличие от жабр отделены от окружающей среды стенками тела, тканями и внеклеточной жидкостью, и поэтому они могут контролировать состав внутренней среды организма. Хотя все рыбы живут в водной среде, механизм экскреции и осморегуляции у пресноводных и морских рыб настолько различен, что эти две группы следует рассмотреть по отдельности.
Пресноводные рыбы. У пресноводных костистых рыб осмолярность жидкостей тела составляет около 300 мосмоль/л, и они гипертоничны по отношению к внешней среде. Несмотря на относительную непроницаемость наружного покрова из чешуй, покрытых слизью, внутрь тела путем осмоса поступает значительное количество воды через высокопроницаемые жабры, и через них же теряются соли. Жабры служат также органами выделения таких азотистых продуктов, как аммиак. Для поддержания стационарного состояния жидкостей внутренней среды пресноводные рыбы должны постоянно выводить много воды. Это происходит у них благодаря образованию большого объема ультрафильтрата, из которого растворенные вещества извлекаются путем их избирательной реабсорбции в капилляры, окружающие почечные канальцы. Почки образуют большое количество сильно разведенной мочи (гипотоничной по отношению к крови), в которой содержится и ряд других растворенных веществ. Количество мочи, выделяемой за сутки, может составлять до одной трети от всей массы тела. Потеря солей с мочой возмещается за счет электролитов, получаемых с пищей, и за счет активного поглощения их из окружающей воды особыми клетками, находящимися в жабрах.
Морские рыбы. Полагают, что рыбы впервые появились в морской среде, затем успешно заселили пресноводные водоемы, и после этого некоторые из них вторично переселились в море, дав начало пластиножаберным и морским костистым рыбам. В процессе эволюции в пресной воде у рыб сформировались многие физиологические механизмы, приспособленные к такому осмотическому давлению жидкостей тела, которое в 2-3 раза меньше, чем у морской воды. После возвращения рыб в морскую среду жидкости их тела сохранили осмотическое давление, присущее их предкам, и в связи с этим возникла проблема гомеостаза жидкостей тела в условиях гипертонического окружения (рис. 19.14).
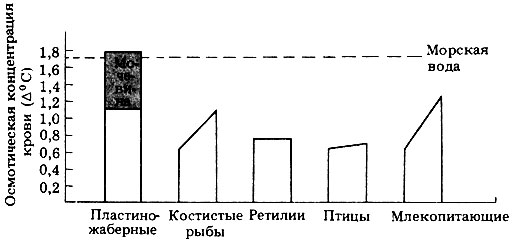
Рис. 19.14. Приблизительные концентрации растворенных веществ в жидкостях тела морских позвоночных. Пластиножаберные рыбы - единственные позвоночные, у которых жидкости тела гипертоничны по отношению к окружающей среде; но, как видно из диаграммы, концентрация электролитов у них лишь ненамного выше, чем у костистых рыб. Благодаря задержке мочевины осмотическое давление у них такое же, как у морской воды, о чем свидетельствует депрессия точки замерзания (Δ°С)
Пластиножаберные рыбы. У этих рыб исходная осмолярность жидкостей тела примерно такова же, что и у морских костистых рыб, т.е. эквивалентна 1%-ному раствору соли. Избыточная потеря воды в гипертонической морской воде предотвращается за счет синтеза и удержания мочевины в тканях и жидкостях тела. По-видимому, большинство клеток тела, за исключением клеток головного мозга, способно синтезировать мочевину, и для своей метаболической активности они не только нуждаются в присутствии мочевины, но и обладают толерантностью к высоким ее концентрациям. Исследования, проведенные на изолированных сердцах акул, показали, что сердце может сокращаться только при перфузии сбалансированным солевым раствором, содержащим мочевину. Жидкости тела у акул содержат 2-2,5% мочевины, что в 100 раз превышает концентрацию, переносимую другими позвоночными. Как правило, высокая концентрация мочевины приводит к разрыву водородных связей, денатурации белков и тем самым к инактивации ферментов. Однако у пластиножаберных рыб этого почему-то не происходит. Мочевина в сочетании с неорганическими ионами и другим азотистым продуктом обмена - триметиламиноксидом (CH3)3N=0, менее токсичным, чем аммиак,- создают в жидкостях тела более высокое осмотическое давление, чем в морской воде (Δ морской воды составляет 1,7°С, а у жидкостей тела пластиножаберных-1,8°С) (рис. 19.14). Будучи слегка гипертоничными по отношению к окружающей среде, пластиножаберные рыбы поглощают воду путем осмоса через жабры. Вода вместе с избытком мочевины и триметиламиноксидом выводится почками с мочой, которая слегка гипотонична по отношению к жидкостям тела. Почки имеют длинные канальцы, которые используются для избирательной реабсорбции мочевины, а не для выведения поступающих с пищей солей. Избыток же ионов натрия и хлора удаляется из жидкой среды организма путем активной секреции в прямую кишку клетками ректальной железы - маленькой железки, связанной протоком с прямой кишкой. Жабры относительно непроницаемы для отходов азотистого обмена, и их выведение целиком контролируется почками. Таким путем осмотическое давление жидкостей тела поддерживается на высоком уровне.
Морские костистые рыбы. У морских костистых рыб осмотическое давление жидкостей тела поддерживается на уровне более низком, чем у морской воды (рис. 19.14). Благодаря чешуе и слизи наружные покровы рыб относительно мало проницаемы для воды и ионов, но вода легко теряется из организма (а ионы поглощаются) через жабры. Для регуляции состава жидкостей тела костистые рыбы пьют морскую воду, а особые секреторные клетки в кишечнике извлекают из нее доли путем активного транспорта и выделяют их в кровь. В жабрах имеются хлоридные клетки, которые активно поглощают из крови ионы хлора и выделяют их в окружающую среду, а вслед за ионами хлора по принципу сохранения электрохимической нейтральности выходят и ионы натрия. Другие ионы, присутствующие в морской воде в большом количестве,- ионы магния и сульфата - удаляются с изотонической мочой, образуемой в небольшом количестве почками. Почки не имеют клубочков и поэтому не способны к ультрафильтрации. Все компоненты мочи, такие как азотистое соединение триметиламиноксид (придающий рыбе характерный запах) и соли, секретируются в почечные канальцы, а за ними осмотическим путем следует и вода.
Эвригалинные рыбы. Существует ряд видов эвригалинных рыб, которые не только переносят небольшие изменения солености воды, но и могут полностью адаптироваться к жизни в пресной и в морской воде на протяжении длительных периодов их жизни. В зависимости от того, куда эти рыбы движутся на нерест, различают анадромных и катадромных рыб. Анадромные рыбы (греч. аnа - вверх, dromein - бежать), такие как лосось (Salmo salar), выводятся из икры в пресной воде и мигрируют в море; здесь они достигают зрелости и затем возвращаются для нереста в реки. Катадромные рыбы (греч. cata-вниз), к которым относится угорь (Anguilla vulgaris), мигрируют в противоположном направлении. Они выводятся в морской воде и мигрируют в пресноводные водоемы, где достигают зрелости, после чего возвращаются для нереста в море. При переходе из реки в море угорь теряет за 10 часов около 40% своего веса. Чтобы компенсировать эту потерю и сохранить гипотоничность жидкостей тела, он пьет морскую воду, а соли выделяет путем активной секреции через жабры. При переходе угря из моря в реку его масса вначале увеличивается за счет поступления воды путем осмоса, но уже через два дня он достигает стабильного осмотического состояния. В пресной воде угорь поглощает соли через жабры путем активного транспорта.
На примере этих двух групп рыб мы видим, что механизмы активного транспорта в жабрах могут действовать в двух направлениях. Связано ли это с переменой направления работы ионных насосов в одних и тех же клетках или с функционированием разных групп клеток, пока неизвестно. Предполагается, что на эти механизмы влияют гормоны, выделяемые гипофизом и корой надпочечников. У рыб обеих групп при переходе в пресную воду существует период "выжидания", позволяющий механизмам осморегуляции приспособиться к новой среде.
Считается, что амфибии произошли от рыбообразных пресноводных предков и унаследовали от них проблемы осморегуляции, связанные с тем, что их кровь гипертонична по отношению к окружающей среде. Кожа лягушек проницаема для воды, и именно через кожу поступает из внешней среды в организм основная масса воды. Избыток поглощаемой организмом воды удаляется путем ультрафильтрации в многочисленных крупных почечных клубочках.
Почки амфибий широко использовались для изучения физиологии этих органов, так как их крупные клубочки расположены близко к поверхности. В эти клубочки и канальцы можно вводить микрошприц и извлекать из них фильтрат для анализа. Таким путем можно определить эффективность ультрафильтрации и избирательной реабсорбции. Амфибии выделяют большое количество очень разбавленной мочи, гипотоничной по отношению к жидкостям тела. Моча содержит мочевину, которая выводится путем ультрафильтрации и путем секреции в канальцы. Преимущество этого механизма состоит в том, что он позволяет амфибиям снижать скорость клубочковой фильтрации в засушливых условиях и таким образом уменьшать потерю воды с мочой, тогда как канальцы продолжают получать кровь из воротных сосудов почек, из которых в канальцы активно секретируется мочевина. В этом отношении данный механизм противоположен тому, что имеет место у пластиножаберных рыб, у которых мочевина в канальцах активно реабсорбируется.
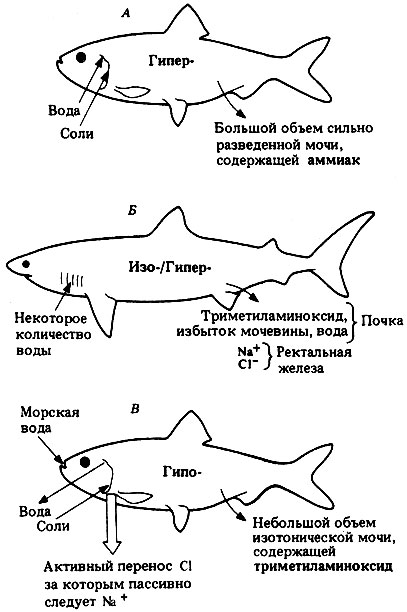
Рис. 19.15. Экскреция и осморегуляция у пресноводных костистых рыб (А), пластиножаберных (Б) и морских костистых рыб (В). Сокращения Гипо-, Изо- и Гипер- указывают тоничность внутренней среды по отношению к внешней
Некоторая часть солей неизбежно теряется с мочой и в результате диффузии через кожу, но эта потеря возмещается за счет солей, поступающих с пищей, а также активно поглощаемых из окружающей воды кожей, которая служит у амфибий главным органом осморегуляции. Личинка бесхвостой амфибии - головастик - является полностью водным организмом и выделяет аммиак через жабры, но при метаморфозе состав азотистых экскретов и механизм их выделения меняются и становятся такими, как описано выше.
Лягушки способны накапливать воду в мочевом пузыре и многочисленных подкожных лимфатических пространствах. За счет этих запасов возмещается потеря воды путем испарения в те периоды, когда лягушка находится на суше. Жабы способны находиться в сухих условиях более продолжительное время, так как их почки могут реабсорбировать воду из клубочкового фильтрата и образовывать более концентрированную мочу, а кожа менее проницаема для воды. Известно, что проницаемость кожи у амфибий регулируется антидиуретическим гормоном, выделяемым задней долей гипофиза; как полагают, механизм регуляции проницаемости здесь тот же, что и в почечных канальцах млекопитающих.
Для нормального функционирования клеток в организме животного необходимо стационарное состояние внутриклеточной жидкости. Гомеостатический обмен водой между клетками, тканевой жидкостью, лимфой, плазмой крови и окружающей средой представляет проблему и для водных, и для наземных организмов. Водные формы получают или теряют воду путем осмоса через все проницаемые участки поверхности тела в зависимости от того, каково окружение - гипотоническое оно или гипертоническое. Наземные организмы сталкиваются с проблемой потери воды и для поддержания устойчивого водного баланса используют многочисленные приспособления, приведенные в табл. 19.5. Это устойчивое состояние водного обмена достигается за счет баланса между отдачей воды и ее получением (табл. 19.6).

Таблица 19.5. Механизмы сохранения воды у наземных животных

Таблица 19.6. Компоненты водного баланса у наземных организмов
Эти животные первыми приспособились к наземной жизни. Они обладают множеством морфологических, биохимических и физиологических адаптаций для существования на суше. Однако во всех трех отрядах (черепахи, ящерицы и змеи, крокодилы) имеются виды, которые вторично приспособились к жизни в пресной и морской воде. У всех этих животных механизмы выделения и осморегуляции адаптированы к соответствующим условиям.
У наземных пресмыкающихся потере воды препятствует относительно непроницаемая кожа, покрытая роговыми чешуйками. Органами газообмена у них служат легкие, расположенные внутри тела, что уменьшает потерю воды. В тканях образуется нерастворимая мочевая кислота, которая может выводиться без большой потери влаги. Для удаления избытка ионов натрия и калия нужна вода, но поскольку экономия воды имеет жизненно важное значение, эти ионы соединяются с мочевой кислотой, образуя нерастворимые ураты натрия и калия, которые удаляются вместе с мочевой кислотой. Почечные клубочки имеют малые размеры и образуют лишь такое количество фильтрата, которое необходимо для вымывания мочевой кислоты из почечных канальцев в клоаку, где часть воды реабсорбируется. У многих наземных рептилий почечные клубочки вообще отсутствуют.
У сухопутных рептилий нет специальных механизмов для выведения солей, а ткани способны переносить повышение концентрации солей на 50% по сравнению с обычным уровнем после приема их с пищей или избыточной потери воды. Морские пресмыкающиеся, такие как галапагосская игуана и зеленая черепаха (Chelone mydas), получают с пищей большое количество соли. Их почки не способны справиться с быстрым выведением этого избытка соли из жидкостей тела, и им помогают особые солевые железы, расположенные на голове. Эти железы способны секретировать раствор хлористого натрия, в несколько раз более концентрированный, чем морская вода. Солевые железы находятся у черепахи в глазницах, и протоки от них идут к глазам; отсюда впечатление, что черепаха плачет. В "слезах", выделяемых солевыми железами черепах, концентрация солей очень высока.
Важной особенностью рептилий и птиц, благодаря которой они могут существовать вне воды в течение всего жизненного цикла, является наличие у них клейдоических яиц (рис. 20.52). Яйцо заключено в плотную оболочку, которая предохраняет зародыша от обезвоживания. В процессе эмбриогенеза вырост задней кишки образует мешковидную структуру, называемую аллантоисом, в которой откладывается мочевая кислота, выделяемая эмбрионом. Поскольку мочевая кислота нерастворима и нетоксична, это служит для эмбриона идеальным способом депонирования экскретов. На более поздних стадиях развития аллантоис васкуляризуется, прижимается к оболочке и функционирует как орган газообмена.
Птицы, по-видимому, произошли от наземных пресмыкающихся, таких как змеи и ящерицы, и унаследовали те же проблемы. Кожа птиц относительно непроницаема для воды, и благодаря наличию перьев и отсутствию потовых желез скорость испарения влаги у птиц очень мала. Однако значительное количество воды теряется у них в дыхательных путях в связи с очень активной вентиляцией легких и сравнительно высокой температурой тела. Вследствие большой интенсивности метаболизма некоторые мелкие птицы могут терять за сутки до 35% веса тела.
Азотистые продукты обмена удаляются в виде мочевой кислоты с мочой, гипертоничной по отношению к жидкостям тела. Моча поступает в клоаку, где часть воды из мочи и фекальных масс всасывается обратно, благодаря чему из организма выводятся почти твердые экскременты.
Почки птиц содержат мелкие клубочки. Вся кровь, снабжающая каналец, в котором происходит реабсорбция воды и секреция солей, поступает от клубочка, для эффективной работы которого необходимо относительно высокое кровяное давление. Таким образом осуществляется связь между образованием большого объема клубочкового фильтрата и последующим всасыванием большой части содержащихся в нем воды и солей. Это всасывание облегчается тем, что поверхность канальца увеличена за счет образования петли Генле. В результате деятельности этой структуры концентрация мочевой кислоты в моче достигает 21%, что почти в 3000 раз выше ее концентрации в жидкостях тела.
Некоторые морские птицы (пингвины, олуши, бакланы, альбатросы), которые питаются рыбой и пьют морскую воду, поглощают большие количества солей. Соли выводятся из жидкостей тела специализированными секреторными клетками солевых, или носовых, желез. Эти железы сходны с солевыми железами морских рептилий и тоже расположены в глазницах. Они выделяют раствор хлористого натрия, концентрация которого в 4 раза выше, чем в жидкостях тела. Носовые железы состоят из множества долек, содержащих большое число секреторных трубочек, которые открываются в центральный проток; этот проток ведет в носовую полость, где раствор соли освобождается в виде больших капель или выдувается в виде мельчайших брызг.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'