21.6.4. Первичный рост побега
Строение типичной апикальной меристемы побега представлено на рис. 21.17 и 21.18. На рис. 21.18 показано примерное подразделение верхушки побега на зоны деления, роста и дифференцировки клеток. У побега эти зоны перекрываются сильнее, чем в корне. В направлении от куполообразной апикальной меристемы назад клетки становятся все старше, так что в одной верхушке побега можно одновременно видеть разные этапы роста. Поэтому изучать последовательные стадии развития растительной ткани относительно несложно.
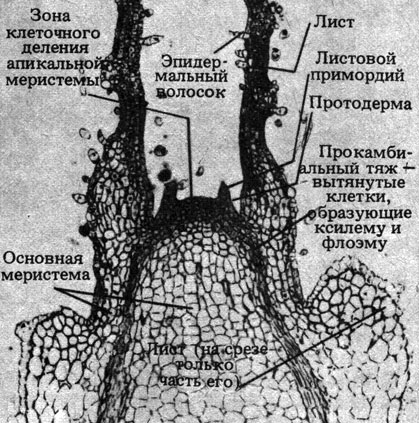
Рис. 21.17. Апикальная меристема
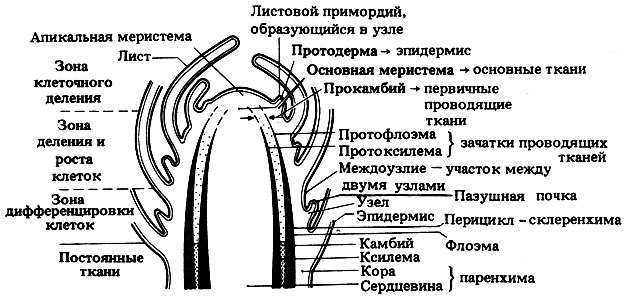
Рис. 21.18. Кончик побега двудольного растения (в продольном разрезе, схема). Видны апикальная меристема и зоны первичного роста. Для простоты проводящие ткани, идущие в листья и почки, не показаны
В побегах различают три основные зоны меристематической ткани: 1) протодерму, дающую начало эпидермису; 2) прокамбий, образующий проводящие ткани, т.е. перицикл, флоэму, васкулярный камбий и ксилему; 3) основную меристему, образующую паренхимные основные ткани (у двудольных это кора и сердцевина). Эти меристематические ткани закладываются в результате деления инициалей в верхушке побега. В зоне роста дочерние клетки, образующиеся путем деления инициалей, увеличиваются в размерах - главным образом за счет осмотического поглощения воды, поступающей в цитоплазму, а из нее - в вакуоли. Рост стеблей и корней в длину достигается в основном за счет происходящего на этой стадии удлинения клеток (рис. 21.19).
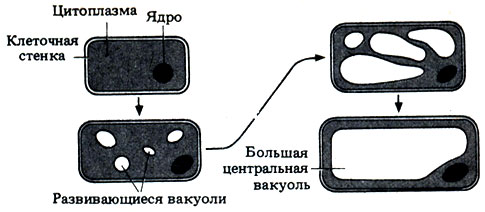
Рис. 21.19. Стадия растяжения в росте меристематической клетки
Мелкие вакуоли увеличиваются в размерах и сливаются в конце концов в одну большую вакуоль. Развивающееся в клетке тургорное давление растягивает ее тонкие стенки, а относительное расположение целлюлозных фибрилл в них определяет окончательную форму клетки. Объем цитоплазмы на конечном этапе может быть лишь немногим больше, чем в исходной меристематической клетке, но теперь вакуоль оттесняет ее на периферию. Когда рост почти завершается, у многих клеток происходит дополнительное утолщение стенок за счет целлюлозы или лигнина, в зависимости от типа клетки. Это может ограничить дальнейший рост, но не обязательно ведет к его полному прекращению. Например, удлинение колленхимных клеток в коре может продолжаться, в то время как на внутренней стороне их первичных стенок откладывается целлюлоза в виде столбиков. Таким образом, клетки колленхимы еще в период своего роста могут нести опорные функции. В отличие от этого на стенках развивающихся клеток склеренхимы откладываются толстые слои лигнина, и клетки вскоре отмирают, т.е. их дифференцировка начинается лишь после того, как рост фактически завершен.
Прокамбий образует ряд продольных тяжей, состоящих из более узких и длинных клеток, чем основная меристема. Первыми в прокамбии начинают дифференцироваться клетки протоксилемы во внутренней его части и клетки протофлоэмы - в наружной. Это элементы первичной ксилемы и первичной флоэмы, которые образуются до завершения роста клеток в длину. В протоксилеме лигнин, как правило, образует только кольцевидные или спиральные утолщения на трахеидах (см. разд. 8.2.1), которые благодаря своей прерывистости делают возможным рост и растяжение целлюлозы между утолщениями по мере роста в длину окружающей ткани. Элементы как протоксилемы, так и протофлоэмы вскоре отмирают и по мере роста окружающих тканей сминаются, растягиваются и наконец спадаются. Их функции принимают на себя ксилема и флоэма, развивающиеся позднее в зоне дифференцировки.
В зоне дифференцировки каждая клетка полностью специализируется для выполнения своей собственной особой функции в соответствии со своим положением в данном органе относительно других клеток. Самые крупные изменения происходят в протокамбиальных тяжах, которые дифференцируются в проводящие пучки. Это связано с лигнификацией стенок склеренхимных волокон и элементов ксилемы, а также с развитием трубок, характерных для сосудов ксилемы и ситовидных трубок флоэмы. Эти ткани в их окончательном виде описаны в разд. 8.2. Теперь склеренхима и ксилема тоже несут опорные функции, выполнявшиеся ранее только колленхимой и тургесцентной паренхимой. Между ксилемой и флоэмой имеются клетки, сохраняющие способность к делению. Они образуют васкулярный камбий, деятельность которого будет рассмотрена позже, при описании вторичного утолщения.
Развитие побега включает также рост листьев и боковых почек. Листья закладываются в виде мелких вздутий или складок, называемых листовыми зачатками; они хорошо видны на рис. 21.17. Эти вздутия состоят из групп меристематических клеток и распределены с равномерными интервалами; места их возникновения называются узлами, а промежутки между ними - междоузлиями. Способ расположения листьев может быть различным и носит название филлотаксиса. Листья могут располагаться мутовками - от каждого узла отходит два или большее число листьев; или же от каждого узла отходит по одному листу, причем листья либо образуют два ряда, направленные в противоположные стороны, либо расположены по спирали. Как правило, они обычно располагаются таким образом, чтобы перекрывание между ними, а значит, и взаимное затенение было минимальным (мозаичное расположение).
Листовые зачатки быстро удлиняются, так что вскоре они окружают апикальную меристему, защищая ее как чисто механически, так и тем теплом, которое они выделяют при дыхании. Позднее они растут, площадь их увеличивается и они образуют листовые пластинки. Деление клеток постепенно прекращается, хотя может продолжаться до тех пор, пока листья не достигнут половины своей окончательной величины.
Вскоре после того, как листья начинают расти, в пазухах между ними и стеблем образуются почки. Они представляют собой небольшие группы меристематических клеток, обычно пребывающих в состоянии покоя, но способных приступить к делению и росту на более поздней стадии. Из этих почек могут развиваться ветки или такие специализированные органы, как цветки, а также подземные структуры-корневища и клубни. Полагают, что пазушные почки находятся под контролем апикальной меристемы (см. разд. 15.3.3 об апикальном доминировании).
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'