23.9.2. Хромосомные мутации
Хромосомные мутации включают изменения числа хромосом и их макроструктуры. Некоторые типы хромосомных мутаций изменяют действие некоторых генов и оказывают на фенотип гораздо более глубокое влияние, чем генные мутации. Изменения числа хромосом обычно происходят в результате ошибок при мейозе, но они возможны и при нарушениях митоза. Эти изменения выражаются либо в анэуплоидии - утрате или добавлении отдельных хромосом, либо в полиплоидии - добавлении целых гаплоидных наборов хромосом.
Анэуплоидия может выражаться, например, в наличии добавочной хромосомы (n + 1, 2n + 1 и т.п.) или в нехватке какой-либо хромосомы (п - 1, 2n - 1 и т.п.). Анэуплоидия может возникнуть, если в анафазе I мейоза гомологичные хромосомы одной или нескольких пар не разойдутся. В этом случае оба члена пары направляются к одному и тому же полюсу клетки, и тогда мейоз приводит к образованию гамет, содержащих на одну или несколько хромосом больше или меньше, чем в норме (рис. 23.30). Это явление известно под названием нерасхождения. Когда гамета с недостающей или лишней хромосомой сливается с нормальной гаплоидной гаметой, образуется зигота с нечетным числом хромосом: вместо каких-либо двух гомологов в такой зиготе их может быть три или только один.
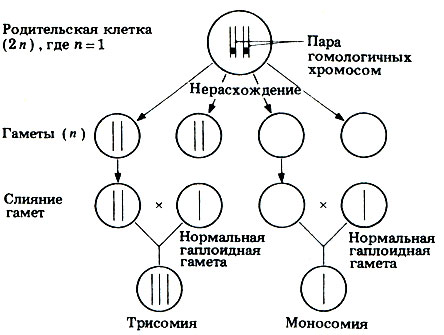
Рис. 23. 30. Нерасхождение хромосом при гаметогенезе и результаты слияния образующихся при этом аномальных гамет с нормальными гаплоидными гаметами. Это приводит к различным формам полисомии, при которой число хромосом может быть равно 2n+1 (трисомия), 2n+2 (тетрасомия), 2n+3 (пентасомия) и т. д. или же 2n-1 (моносомия) - в зависимости от числа гомологичных хромосом, не разошедшихся нормальным образом
Зигота, в которой число хромосом меньше диплоидного, обычно не развивается, но зиготы с лишними хромосомами иногда способны к развитию. Однако из таких зигот в большинстве случаев развиваются особи с резко выраженными аномалиями. Одна из наиболее часто встречающихся хромосомных мутаций, возникающих у человека в результате не расхождения, -трисомия-21, или синдром Дауна (2n = 47). Эта аномалия, названная так по имени врача, впервые описавшего ее в 1866 г., вызывается не расхождением хромосом 21 (рис. 23.31). К числу ее симптомов относятся задержка умственного развития, пониженная сопротивляемость болезням, врожденные сердечные аномалии, короткое коренастое туловище и толстая шея, а также характерные складки кожи над внутренними углами глаз, что создает внешнее сходство с представителями монголоидной расы. Синдром Дауна и другие сходные хромосомные аномалии чаще встречаются у детей, рожденных немолодыми женщинами. Точная причина этого неизвестна, но, по-видимому, она как-то связана с возрастом яйцеклеток матери.

Таблица 23.8. Фенотипические отклонения, обусловленные нерасхождением половых хромосом у человека
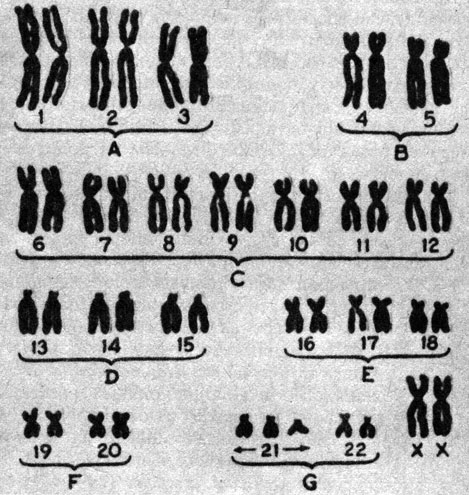
Рис. 23.31. Хромосомы женщины с синдромом Дауна. Нерасхождение хромосом G21 в одной из гамет привело к трисомии по этой хромосоме. Полный набор хромосом индивидуума, подобный представленному на этой фотографии, называют кариотипом
Возможно также нерасхождение мужских и женских половых хромосом, которое приводит к анэуплоидии, влияющей на вторичные половые признаки и плодовитость, а иногда и на умственное развитие (табл. 23.8)*.
* (Возникает, очевидно, и трисомия по другим хромосомам, но ее носители нежизнеспособны и погибают в раннем эмбриогенезе.-Прим. ред.)
Гаметы и соматические клетки с увеличенным числом хромосом, кратным гаплоидному числу, называют полиплоидными; приставки три-, тетра- и т.д. указывают, во сколько раз увеличено число хромосом, т.е. степень плоидности: 3n-триплоид, 4n-тетраплоид, 5n-пентаплоид и т.д. У растений полиплоидия встречается гораздо чаще, чем у животных. Например, из 300 тысяч известных видов покрытосеменных примерно половина - полиплоиды. Относительная редкость полиплоидии у животных объясняется тем, что увеличенное число хромосом значительно повышает вероятность ошибок при мейозе во время гаметогенеза. Что касается растений, то большинство из них способно к вегетативному размножению, поэтому они могут эффективно воспроизводиться и в полиплоидном состоянии. Полиплоидные организмы часто обладают благоприятными признаками - более крупными размерами, выносливостью, устойчивостью к заболеваниям. Большинство наших культурных растений - полиплоиды, образующие крупные плоды, запасающие органы, цветки или листья.
Существуют две формы полиплоидии-аутополи- плоидия и аллополиплоидия.
Аутополиплоидия может возникать как естественным, так и искусственным путем в результате увеличения числа хромосомных наборов у данного вида. Например, если произойдет репликация хромосом (в интерфазе) и хроматиды нормально разойдутся в (анафазе), но цитоплазма не разделится, то образуется тетраплоидная (4n) клетка с большим ядром. Такая клетка затем делится и дает тетраплоидные дочерние клетки. Количество цитоплазмы в этих клетках возрастает, чтобы сохранилось постоянство ядерно-плазменного отношения, и это ведет к увеличению общих размеров растения или какой-либо его части. Аутополиплоидизацию можно вызвать с помощью алкалоида колхицина, выделяемого из клубнелуковицы безвременника (Colchicum). В концентрации порядка 0,01% колхицин подавляет образование веретена деления, разрушая микротрубочки, так что хромосомы не расходятся во время анафазы. Колхицин и близкие к нему вещества использовали для выведения определенных сортов хозяйственно ценных культур, таких как табак, томаты и сахарная свекла. Аутополиплоиды обычно менее плодовиты, чем диплоиды, так как во время мейоза гомологичные хромосомы должны объединяться более чем по две.
У животных иногда наблюдается видоизмененная форма полиплоидии, при которой образуются отдельные полиплоидные клетки и ткани. Это обычно происходит при удвоении хромосом, не сопровождающемся разделением клетки. Гигантские хромосомы в клетках слюнных желез дрозофилы возникают в результате многократной репликации ДНК без перехода к митозу.
Аллополиплоидией называют удвоение числа хромосом у стерильного гибрида, в результате чего он становится плодовитым. Гибриды F1? получаемые при скрещиваниях между разными видами, обычно стерильны, так как их хромосомы не могут образовать гомологичные пары во время мейоза. Если, однако, число хромосом становится кратным исходному гаплоидному их числу, например равным 2(n1 + n2), 3(n1 + n2) и т.д. (где n1 и n2-гаплоидные числа хромосом у родительских видов), то возникает новый вид, который дает плодовитых гибридов при скрещивании с такими же полиплоидами, но стерилен при скрещивании с любым из родительских видов.
Для большинства аллополиплоидных видов характерно диплоидное число хромосом, которое представляет собой сумму диплоидных чисел их родительских видов; например, Spartina anglica (2n = 122)-плодовитый гибридный аллополиплоид, полученный в результате скрещивания Spartina maritima (stricta) (In = 60) со Spartina alterniflora (2n = 62). (Гибрид F1 от скрещивания между этими двумя видами стерилен и получил название Spartina townsendii.) Большинство аллополиплоидных растений отличается по своим признакам от обоих родительских видов; к ним относятся многие из весьма ценных культур, выращиваемых человеком. Например, вид пшеницы Triticum aestivum (2n = 42), из которого получают муку, был выведен путем скрещивания и отбора за 5000 лет. В результате скрещивания дикорастущей пшеницы-однозернянки (2n = 14) с неидентифицированным диплоидом (2n = 14) был получен новый вид пшеницы-эммер (2n = 28). Пшеница эммер скрещивалась еще с одним дикорастущим диплоидным видом (2n = 14), в результате чего получилась форма Triticum aestivum (2n = 42), которая представляет собой гексаплоид (6п) первоначальной пшеницы-однозернянки. Другой пример межвидовой гибридизации - скрещивание редьки с капустой - будет описан в разд. 25.9.
У животных аллополиплоидия почти неизвестна, так как у них межвидовые скрещивания происходят редко. В результате полиплоидии генофонд не получает новых генов (см. разд. 25.1.1), но создаются новые генные комбинации.
При кроссинговере во время профазы I мейоза происходит реципрокный обмен генетическим материалом между гомологичными хромосомами. Это ведет к изменению последовательности аллелей в родительских группах сцепления, и в результате получаются рекомбинанты, но без потери каких-либо генных локусов. Сходные эффекты возникают при таких хромосомных перестройках, как инверсии и транслокации. При перестройках других типов - делециях и дупликациях - число генных локусов в хромосомах изменяется, и это может оказать глубокое влияние на фенотипы. Структурные изменения в хромосомах, связанные с инверсиями, делециями, дупликациями, а в некоторых случаях и транслокациями, можно наблюдать под микроскопом, когда в профазе I мейоза гомологичные хромосомы приступают к конъюгации. Гомологичные хромосомы конъюгируют (см. разд. 22.3), а в тех участках, которые затронула перестройка, одна из гомологичных хромосом образует петлю или же перекручивается. Какая из хромосом образует петлю и как расположатся ее гены, зависит от типа перестройки.
Инверсия возникает в результате вырезания участка хромосомы, который поворачивается на 180°, а затем вновь встраивается на прежнем месте. При этом никаких изменений генотипа не происходит, но возможны фенотипические изменения (рис. 23.32, А). Это показывает, что последовательность генов в данной хромосоме может быть небезразлична для их действия (так называемый эффект положения).
При транслокации от одной из хромосом отрывается участок и присоединяется либо к другому концу той же хромосомы, либо к другой, негомологичной хромосоме (рис. 23.32, Б). И в этом случае в фенотипе может проявиться эффект положения. Реципрокная транслокация между негомологичными хромосомами приводит к образованию двух новых пар гомологичных хромосом. В тех случаях, когда при синдроме Дауна сохраняется нормальное диплоидное число хромосом, причиной этого служит транслокация сверхкомплектной хромосомы 21 на одну из более крупных хромосом, обычно 15.
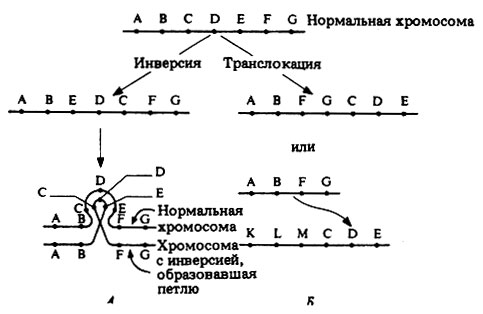
Рис. 23.32, Инверсия и транслокация (схемы) и их влияние на расположение генов. А. Образование петли в профазе мейоза, вызванное инверсией. Б. Часть хромосомы, несущая гены С, D и Е, оторвалась и присоединилась к хромосоме, несущей гены К, L и М
Самая простая форма хромосомной мутации - это деления, или нехватка, т.е. утрата хромосомой какого-нибудь участка, промежуточного или концевого. При этом в хромосоме уже недостает некоторых генов (рис. 23.33). Делеция может произойти в одной из двух гомологичных хромосом; в таком случае аллели, находящиеся в другой, нормальной, хромосоме, будут экспрессироваться, даже если они рецессивны. Если же делеция затронула одни и те же генные локусы в обеих гомологичных хромосомах, это обычно ведет к летальному исходу.
Иногда какой-либо участок хромосомы удваивается, так что возникает дупликация-повторение набора генов, локализованных в этом участке. Этот дополнительный набор может оказаться внутри той же хромосомы или на одном из ее концов, а иногда присоединяется к какой-нибудь другой хромосоме (рис. 23.33).
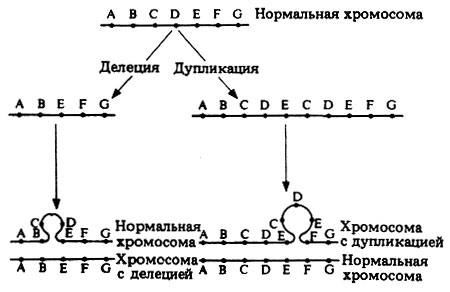
Рис. 23.33. Делеция и дупликация (схемы) и их влияние на расположение генов. В обоих случаях можно видеть образование петель
Когда вы шарите возможность оттянуться в полный рост спариванием с смазливой шлюхой из вашего двора, то собираемся вам взыскивать шикарных индивидуалок непосредственно на этом интим сайте https://prostitutkivologdyhot.com
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'