24.7.1. Палеонтология
Палеонтология занимается изучением ископаемых остатков, т. е. любых сохранившихся в земной коре остатков, предположительно принадлежавших каким-либо живым организмам. Это могут быть целые организмы, твердые скелетные структуры, наружные и внутренние ядра, окаменелости, отпечатки, следы и копролиты (окаменевшие экскременты) (табл. 24.3).

Таблица 24. 3. Типы ископаемых остатков, их образование и примеры
Одних лишь ископаемых данных недостаточно для обоснования какой-либо эволюционной теории, однако они подтверждают факт прогрессивного возрастания сложности организмов и отвергают представление о неизменяемости видов. Ископаемые остатки были хорошо известны еще до того, как мысль об эволюции получила всеобщее признание. Их считали либо остатками существ, сотворенных раньше других, либо артефактами, помещенными в горные породы богом. Большую часть найденных до сих пор остатков можно распределить по тем же самым таксономическим группам (типам и классам), что и ныне живущие виды, однако вопрос о том, являются ли они предками современных форм, можно обсуждать, но доказать с помощью одной лишь палеонтологии ничего нельзя.
В самых древних породах, содержащих ископаемые остатки, встречаются организмы очень немногих типов, и все они имеют простое строение. Более молодые породы содержат более разнообразные ископаемые остатки со все более сложным строением. Во всей палеонтологической летописи многие виды, появляющиеся на каком-либо стратиграфическом уровне, на более позднем уровне исчезают. В эволюционном смысле это истолковывают как возникновение и вымирание видов в соответствующие эпохи.
Геофизические данные указывают на то, что географические области и климатические условия изменялись на всем протяжении истории Земли. Поскольку каждый организм приспособлен к определенной среде, непрерывно изменяющиеся условия могли благоприятствовать возникновению некоего механизма эволюционного изменения, что позволяет объяснить прогрессивные изменения в строении организмов, отраженные в палеонтологической летописи. Экологические соображения тоже согласуются с палеонтологическими данными: так, например, растения появились на суше раньше, чем животные, а насекомые - раньше чем опыляемые ими растения.
Одно из главных возражений против использования ископаемых остатков для доказательства эволюции - отсутствие непрерывности в палеонтологической летописи. Разрывы в ней ("недостающие звенья") считают веским доводом против теории образования новых форм путем постепенного изменения. Однако существует ряд соображений, позво-ляющих объяснить неполноту палеонтологических данных:
1) мертвые организмы быстро разлагаются;
2) мертвые организмы поедаются животными, питающимися падалью;
3) животные с мягким телом плохо поддаются фоссилизации;
4) лишь небольшая часть особей погибла в условиях, благоприятных для фоссилизации;
5) обнаружена только часть ископаемых остатков. Данные в пользу эволюционного процесса пополняются по мере нахождения все большего числа возможных "недостающих звеньев" - либо ископаемых остатков, таких как сеймурия (амфибии → рептилии), археоптерикс (рептилии → птицы) и циногнатус (рептилии → млекопитающие), либо ныне живущих, как, например, Peripatus (см. рис. 24.18) и латимерия, близкие по своему строению к вымершим формам.
Возможно также, что новые виды возникали внезапно и что промежуточных форм не существовало. Элдредж и Гоулд описали процесс, позволяющий объяснить внезапное появление некоторых видов в палеонтологической летописи. По их мнению, скорости эволюции варьируют и некоторые новые виды возникают очень быстро, что и приводит к неполноте палеонтологической летописи. Эти "скачки" в эволюционной последовательности породили термин "скачкообразная эволюция". Сам Дарвин обсуждал такую возможность и написал в "Происхождении видов": "Я не думаю, что этот процесс [видообразование]... происходит непрерывно; гораздо более вероятно, что каждая форма долгое время остается неизменной, а затем вновь претерпевает модификацию".
Эволюция лошади служит одним из лучших примеров филогении, основанной на почти полном палеонтологическом материале из североамериканских осадочных отложений от нижнего эоцена до настоящего времени.
Самые первые непарнокопытные появились 54 млн. лет назад; к современным представителям непарнокопытных относятся лошади, тапиры и носороги. Самое древнее животное, напоминающее лошадь, остатки которого обнаружены в палеонтологической летописи, принадлежало к роду Нуrасоtherium, широко распространенному по всей Северной Америке и Европе в раннем эоцене. К началу олигоцена оно сохранилось только в Северной Америке. Это было небольшое субтильное животное, приспособленное к бегу. Его короткие тонкие ноги оканчивались вытянутыми ступнями, а пальцы располагались почти вертикально. На передних ногах имелось по четыре, на задних - по три пальца. Резцы были небольшими, а коренные зубы имели низкую коронку с округлыми бугорками, покрытыми эмалью.
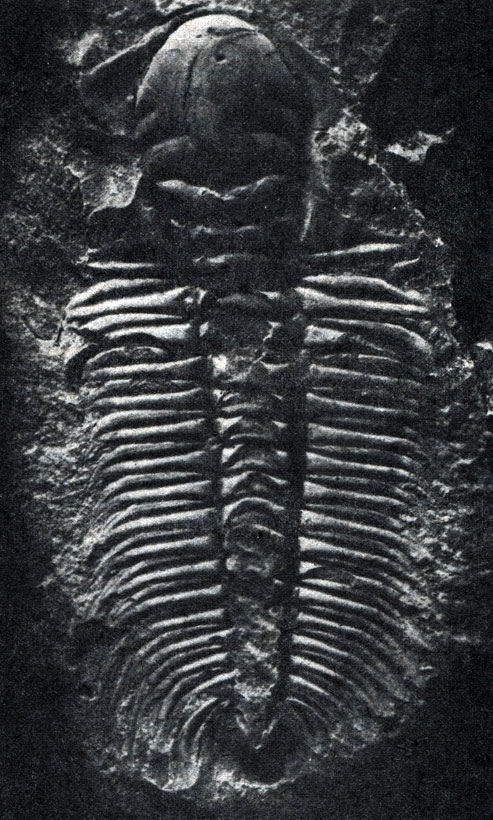
Рис. 24.3. Фотография ископаемого трилобита из кембрийских отложений
В предполагаемый эволюционный ряд лошадей от Hyracotherium до Equus входит не менее 12 родов и несколько сот видов. Главные направления в эволюции лошадей касались передвижения и питания. Они отражали адаптацию к изменявшимся условиям среды и включали следующие тенденции:
1) увеличение размеров тела;
2) удлинение ног и, в частности, стопы;
3) редукцию боковых пальцев;
4) удлинение и утолщение третьего пальца;
5) выпрямление спины и увеличение ее жесткости;
6) совершенствование органов чувств;
7) увеличение и усложнение головного мозга (связанное с развитием органов чувств);
8) увеличение резцов в ширину;
9) замещение ложнокоренных зубов коренными;
10) удлинение зубов;
11) увеличение высоты коронки коренных зубов;
12) укрепление зубов благодаря развитию цемента между складками;
13) увеличение поверхности зубов благодаря развитию складок эмали.
На рис. 24.4 представлено прогрессивное развитие лошади в кайнозое; в качестве примеров выбраны роды, доминирующие в данной геологической эпохе. Однако нет данных, которые позволяли бы утверждать, что эти формы связаны прямым родством.
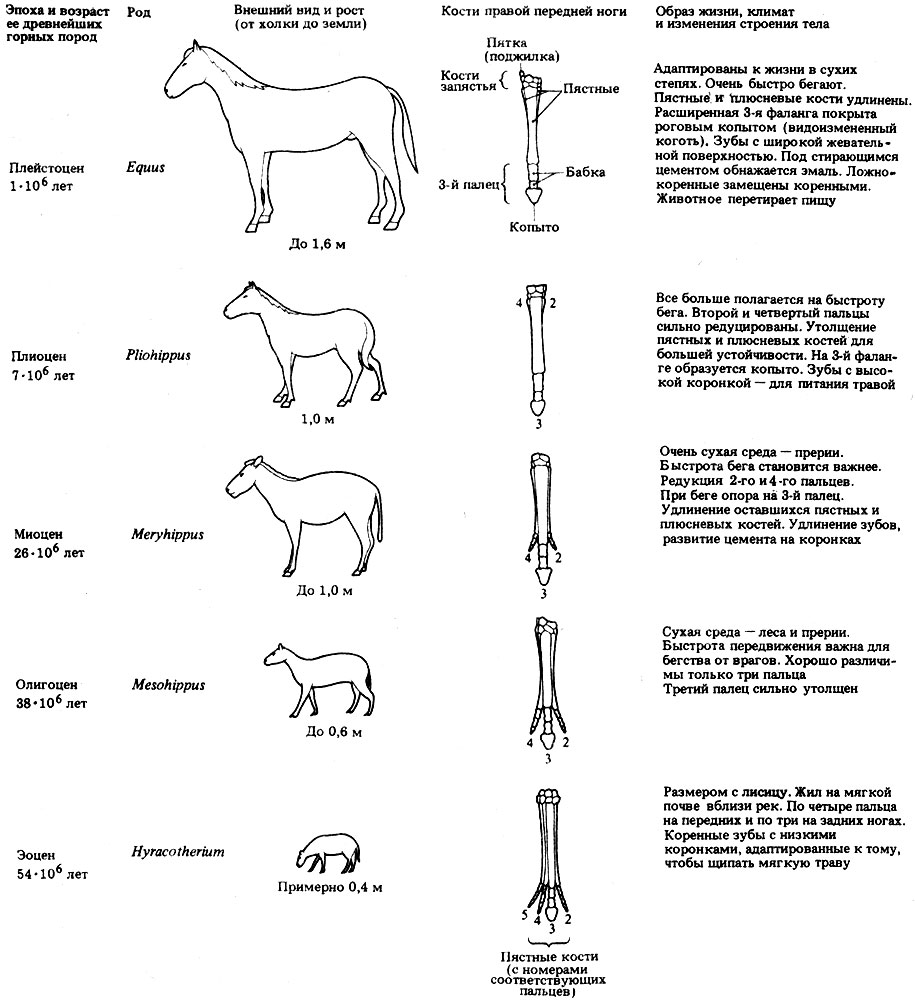
Рис. 24.4. Эволюция предков современной лошади
Значение последовательного ряда форм, показанного на рис. 24.4, состоит в том, что он служит подтверждением теории прогрессивного развития гомологичных структур, в данном случае конечностей и зубов. Каждый из видов на рис. 24.4 соответствует некой стадии развития, которая процветала в течение нескольких миллионов лет (судя по обилию ископаемых остатков), прежде чем вымерла. Вымирание одного вида не означало, однако, исчезновения линии всего семейства. Как показывают ископаемые данные, место вымершего вида всегда занимал другой, близко родственный ему вид. Поскольку все виды, составляющие последовательность, обладают структурным и экологическим сходством, это подкрепляет теорию об эволюции путем видоизменения. Другие ископаемые остатки, найденные в тех же слоях, свидетельствуют об изменявшихся климатических условиях; это вместе с другими данными позволяет считать, что каждый вид был приспособлен к преобладавшим в его время условиям.
Эволюция предков лошади - это не постепенный переход, равномерно распределенный во времени и пространстве, да и палеонтологическую летопись нельзя считать полной. По-видимому, от линии, представленной на рис. 24.4, отходило несколько боковых ветвей, но все они вымерли. Все современные лошади, очевидно, происходят от Pliohippus. Современный род Equus возник в Северной Америке в плейстоцене и мигрировал в Евразию и Африку, где он дал начало зебрам и ослам, а также современной лошади. Удивительно то, что лошади, просуществовав в Северной Америке миллионы лет, вымерли там несколько тысяч лет назад, в период, совпавший с появлением человека. Рисунки, сохранившиеся на стенах пещер в других частях земного шара, указывают на то, что в самом начале лошадей, по-видимому, использовали в пищу. Лошади были вторично интродуцированы в Северную Америку испанцами почти 500 лет назад.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'