15.2.1. Ауксины и фототропизм
Ауксины были открыты в результате изучения фототропизма, которое было начато еще в опытах Чарлза Дарвина и его сына Френсиса. Взяв ко- леоптили овса, они показали на этом очень удобном объекте (рис. 15.3), что рост проростков в сторону света обусловлен тем, что от верхушки стебля передается какое-то "влияние" на лежащую позади зону роста. Часть этих экспериментов представлена на рис. 15.4, где каждая схема отражает результаты не одного, а многих опытов.
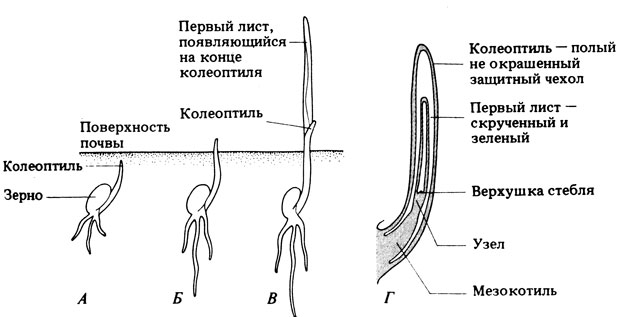
Рис. 15.3. Прорастание типичного проростка злака. А, Б и В - отдель-ные стадии прорастания; Г-разрез колеоптиля на стадии Б
15.6. а) Подробно запишите выводы, которые вы могли бы сделать на основании опытов А-Г на рис. 15.4, учитывая, что изгибы стебля обусловлены изменениями роста в зоне, расположенной ниже верхушки стебля.
б) Почему нужно было поставить опыт В, узнав результаты опыта Б?
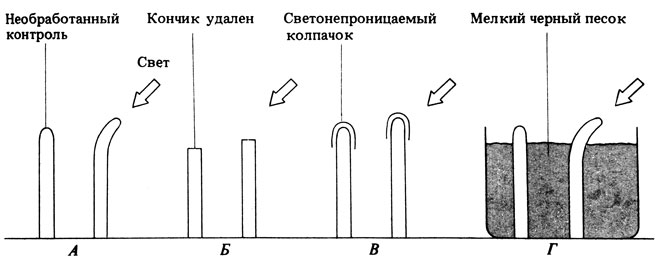
Рис. 15.4. Опыты Дарвина по фототропизму колеоптилей овса. А, Б, В и Г розные опыты; слева в каждом случае показано воздействие, справа - его результат
Если ответную реакцию представить схемой: раздражитель → рецептор → передача сигнала → эффектор → реакция, то оказывается, что меньше всего мы знаем о природе передачи сигнала. В 1913 г. датский физиолог растений Бойсен-Йенсен впервые исследовал этот вопрос. Некоторые из его опытов отображены на рис. 15.5.
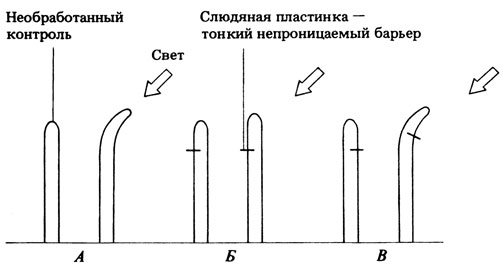
Рис. 15.5. Опыты Бойсен-Йенсена по фототропизму колеоптилей овса. А, Б и В - разные опыты; слева в каждом случае показано воздействие, справа его результат
15.7. Какую дополнительную информацию дали опыты Бойсен-Йенсена?
15.8. Какие результаты будут получены, если повторить эти опыты при равномерном освещении со всех сторон? Нарисуйте схемы, объясните ответ.
В 1928 г. датский физиолог растений Вент окончательно доказал, что существует специфический химический передатчик. Вент поставил перед собой цель перехватить и собрать это вещество в тот момент, когда оно распространяется из верхушки назад, а затем показать его эффективность в различного рода тестах. Он рассуждал, что небольшая диффундирующая молекула должна беспрепятственно проникать внутрь небольшого блока из агарового геля, в котором между молекулами агара остаются довольно большие свободные пространства. Некоторые опыты Вента представлены на рис. 15.6.
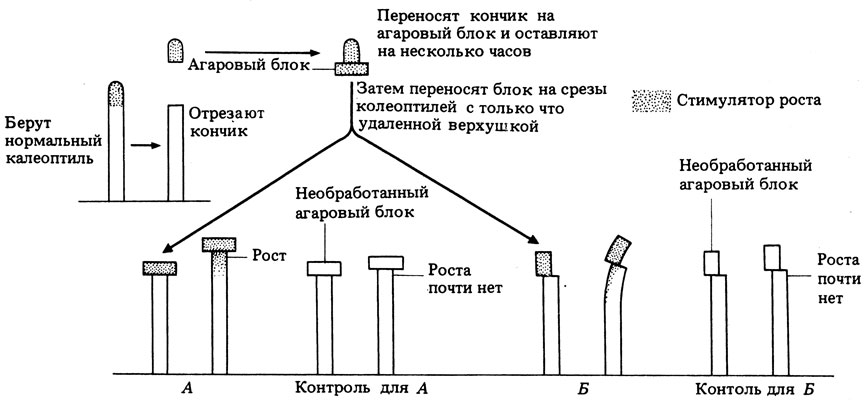
Рис. 15.6. Опыты Вента. А и Б-разные опыты; слева в каждом случае показано воздействие, справа - его результат. Рядом показаны контрольные эксперименты. Все процедуры проводились в темноте или при постоянном освещении
15.9. Какие выводы позволяют сделать результаты, показанные на рис. 15.6?
15.10. Какой результат будет получен, если обработанный блок поместить на правую сторону декапитированного колеоптиля в варианте Б?
На рис. 15.7 представлен еще один эксперимент Вента, который заслуживает особого внимания. В контрольных опытах кончик колеоптиля, помещенный на два агаровых блока А и Б, инкубировали при равномерном освещении или в темноте, а затем переносили агаровые блоки на колеоптиль с удаленным кончиком; величина изгиба, индуцируемого блоками А и Б, в этом случае была одинакова. Одностороннее же освещение верхушки колеоптиля приводило к неравномерному распределению активного вещества между блоками А и Б (рис. 15.7). Это не только подтверждает выводы Бойсен-Йенсена о влиянии света на распределение активного вещества, но и показывает, что можно определять количество этого вещества биологическим методом (с помощью "биотеста"). Вент установил, что величина изгиба колеоптилей овса прямо пропорциональна концентрации активного вещества (в диапазоне ее нормальных физиологических величин).
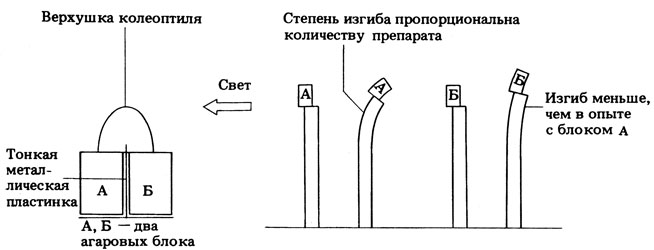
Рис. 15.7. Опыт Вента, демонстрирующий влияние одностороннего освещения на распределение химического фактора (ауксина)
Впоследствии это вещество было названо "ауксином" (от греч. auxein-увеличивать). В 1934 г. оно было идентифицировано как индолилуксусная кислота (ИУК). Вскоре выяснилось, что ИУК широко распространена у растении и что с нею тесно связано увеличение размеров клеток. На рис. 15.8 показано, как, по современным представлениям, передвигается ИУК при одностороннем освещении колеоптилей. Следует, однако, отметить, что колеоптиль - это самая простая из изученных до сих пор систем и что другие системы, по-видимому, устроены гораздо сложнее. Кроме того, нет почти никаких данных о том, что градиент ауксина создается еще во время критического периода, до проявления ответной реакции.
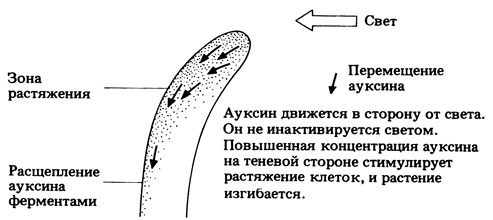
Рис. 15.8. гипотеза, объясняющая влияние одностороннего освещения на распределение ауксина в колеоптиле
Строение ИУК Строение ИУК показано на рис. 15.9.
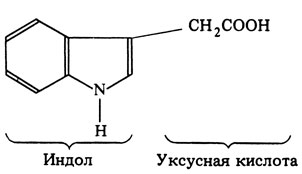
Рис. 15.9. Строение индолилуксусной кислоты
Вскоре были выделены и другие вещества, сходные с ИУК по структуре и активности; удалось также синтезировать несколько соединений подобного типа, так что появился целый класс ростовых веществ, называемых ауксинами. Некоторые из них будут рассмотрены в разд. 15.2.5.
Ауксины непрерывно образуются в точке роста стебля и в молодых листьях. Перемещение ауксинов от верхушки можно назвать базипетальным (от точки роста к основанию органа) и полярным (т.е. идущим в одном направлении). По-видимому, они движутся от клетки к клетке путем диффузии и в конце концов инактивируются и разрушаются ферментами. Транспорт на дальние расстояния про-исходит по проводящей системе (в основном по флоэме) и направлен от побегов к корням. Вероятно, в корнях ауксины почти не образуются. Влияние различных концентраций ауксина на рост побега можно оценить, поставив опыт вроде описанного ниже..
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'