1.4. Функциональное значение клеточной организации
В клетках прокариотов внутриклеточная дифференцировка выражена довольно слабо и определяется факторами, не имеющими отношения к мембранам. Эти клетки не способны заглатывать целиком макромолекулы с помощью пиноцитоза (впячивания плазматической мембраны и образования замкнутых пузырьков внутри клетки). Однако небольшие молекулы могут проникать через плазматическую мембрану в обоих направлениях. Клетки прокариотов обладают также способностью выпускать во внеклеточное пространство некоторые белки. Процесс этот протекает с высокой специфичностью.
Внутреннее пространство в клетках эукариотов разделено на несколько автономных отсеков, причем роль физических границ между этими отсеками играют липопротеидные мембраны. Разнообразные процессы, определяющие функциональную природу клетки, протекают и в отсеках клетки, ограниченных мембранами, и на самих мембранах. Такое разделение внутреннего пространства (компартментализация) весьма выгодно клеткам. Например, эффективность взаимодействия участников отдельных метаболических процессов, субстратов и ферментов резко возрастает, если реакция между ними протекает в небольшом ограниченном объеме. Большое значение имеет также разделение в пространстве прямых и обратных реакций метаболического цикла, а также разделение конкурентных процессов; именно поэтому биосинтез большого числа компонентов клетки и их деградация протекают в различных пространственно обособленных участках клетки. Следует также отметить, что взаимодействие между двумя соседними отсеками клетки может облегчаться благодаря специальным механизмам, регулирующим движение материалов через мембрану, разделяющую эти отсеки. Отмеченные здесь разнообразные аспекты мембранной активности обсуждаются более подробно в гл. 5.
Процессы, определяющие жизнеспособность клеток эукариотов, а также способность их к самовоспроизведению, протекают во внутриядерном пространстве и в цитоплазме (рис. 17). Связь между ядром и цитоплазмой осуществляется через поры, имеющиеся в ядерной мембране. В ядрах содержится дезоксирибонуклеиновая кислота (ДНК), организованная вместе с другими молекулами в хромосомы. Через определенное время происходит репликация ДНК, причем в течение этого процесса ДНК связана с ядерной мембраной. Синтез различных рибонуклеиновых кислот (транспортной, рибосомной и информационной РНК), участвующих в биосинтезе белка, также осуществляется во внутриядерном пространстве. Молекулы синтезированной РНК выходят в цитоплазму через поры в ядерной мембране, причем рибосомная, а возможно, и информационная РНК образуют специфические комплексы с белками. Механизм транспорта рибонуклеиновых кислот через ядерную мембрану неизвестен. Можно лишь предполагать, что во время этого процесса поры не обладают свободной проницаемостью для других молекул.
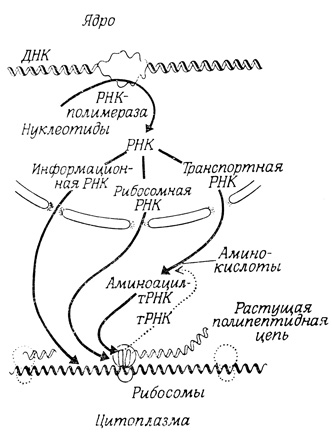
Рис. 17. Схема синтеза полипептидных цепей
Та область цитоплазмы, в которую попадают молекулы РНК, вышедшие из ядра, и в которой полностью проявляется их функциональная активность, является ключевым участком, где осуществляется большая часть характерных для клетки биосинтетических процессов. Именно здесь "растворимые" ферменты катализируют гликолиз - фундаментальный метаболический процесс, в результате которого образуется АТФ, универсальный источник энергии в клетке. Гликолиз и ряд вспомогательных реакций (пентозофосфатный путь) приводят к образованию важных промежуточных продуктов биосинтетических процессов, в том числе НАДФ⋅Н (восстанавливающего агента, принимающего участие в большом числе биосинтетических процессов, в том числе в синтезе жирных кислот и холестерина). Жирные кислоты синтезируются при участии мультиферментных комплексов в цитоплазме. Они входят в состав фосфолипидов, играющих важную роль в мембранных структурах. Включение жирных кислот в фосфолипиды осуществляют ферменты, локализованные в мембранах эндоплазматической сети. На этих мембранах протекают также последние стадии синтеза холестерина, другого важного липидного компонента биологических мембран. Белки синтезируются как на рибосомах, свободно плавающих в цитоплазме, так и на рибосомах, прикрепленных к мембранам эндоплазматической сети и к внешнему слою ядерной мембраны.
Цитоплазма находится в непосредственном контакте с плазматической мембраной, которая играет ключевую роль в осуществлении контроля за движением ионов, питательных веществ и метаболитов из клетки и в клетку. Для большинства веществ плазматическая мембрана служит просто барьером, ограничивающим их движение. Однако для некоторых веществ, имеющих особое значение в метаболизме клетки, таких, как сахара, аминокислоты и некоторые ионы, роль плазматической мембраны более специфична (гл. 2). В этих случаях в мембранах включаются механизмы, облегчающие движение метаболита через клеточную мембрану по градиенту его концентрации. Более сложные механизмы регулируют перенос веществ через мембрану против градиентов концентрации. Это происходит тогда, когда клетки активно накапливают какие-либо вещества, в которых они нуждаются, или выводят те вещества, концентрация которых должна поддерживаться на низком уровне. При таком активном транспорте веществ через мембрану расходуется метаболическая энергия, запасенная в форме АТФ. Плазматическая мембрана выполняет определенную роль в возникновении клеточного ответа на внешние воздействия, например на воздействие гормонов, антигенов, нервных импульсов и т. д.
Гликолиз, являясь центральным метаболическим процессом у организмов, тем не менее делает доступной для использования в клетке лишь небольшую часть энергии, заключенной в углеводных молекулах. Со значительно большим выходом АТФ образуется в цикле трикарбоновых кислот и при окислительном фосфорилировании (рис. 18) -процессах, состоящих из последовательности метаболических реакций, которые протекают в митохондриях всех аэробных (потребляющих кислород) клеток. Пируват, трехуглеродный метаболит глюкозы, образующийся при гликолизе, попадает в митохондрии, где окисляется, превращаясь в CO2 и ацетил-КоА. Последний окисляется с образованием двух новых молекул CO2. Этот процесс состоит из последовательности регулируемых стадий, которые и составляют цикл трикарбоновых кислот. В митохондриях также протекают соответствующие реакции, превращающие различные метаболиты в ацетил-КоА или в промежуточные продукты цикла трикарбоновых кислот, где эти вещества и претерпевают дальнейшее окисление. Важным результатом всех этих процессов является образование НАД⋅Н и ФАД⋅Н - восстановленных коферментов окислительно-восстановительного цикла. Высокая восстанавливающая способность этих соединений используется затем на начальных стадиях транспорта восстановительных эквивалентов по дыхательной цепи (последовательность реакций, в которых с помощью ферментов и коферментов осуществляется перенос электронов), конечные компоненты которой восстанавливают атмосферный кислород. В процессах окислительного фосфорилирования свободная энергия, освобождающаяся при переносе восстановительных эквивалентов по дыхательной цепи, используется для синтеза АТФ. Выход АТФ из одной молекулы глюкозы возрастает благодаря этим процессам с 2 молекул, получаемых при анаэробном гликолизе, до 38.
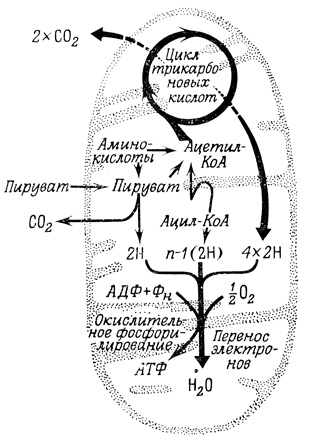
Рис. 18. Цикл трикарбоновых кислот и окислительное фосфорилирование
Ферменты цикла трикарбоновых кислот локализованы в матриксе и на внутренней мембране митохондрий. Ферменты дыхательной цепи и окислительного фосфорилирования образуют мультиферментный комплекс, связанный с внутренней мембраной митохондрий.
В зеленых растениях граны хлоропластов содержат систему, улавливающую энергию из внешней среды. Важным компонентом этой системы является хлорофилл. В результате действия такой системы энергия солнечного света преобразуется в потенциальную энергию восстановительных эквивалентов, а также в энергию АТФ, которая используется затем для восстановления атмосферной двуокиси углерода до органических молекул, таких, как сахара и аминокислоты. Процессы образования различных органических молекул при восстановлении CO2 протекают с участием ферментов, также присутствующих в хлоропластах.
В клетках непрерывно синтезируются различные вещества, предназначенные не только для роста, размножения и экспортных целей, но также и для поддержания нормального существования. Между тем даже в клетках, не проявляющих тенденции к росту или к обильному экспорту продуктов во внешнюю среду, концентрация синтезируемых компонентов не изменяется; следовательно, синтез в таких клетках должен быть полностью уравновешен процессами деградации. Это динамическое равновесие, при котором химический состав системы остается неизменным, тогда как отдельные компоненты непрерывно обновляются, представляет собой нормальный метаболический "круговорот" (рис. 19). При этом клетке необходимо как-то избавляться от поврежденных и непригодных к использованию компонентов. В отдельных случаях организму удается даже избавляться от целых клеток, например при исчезновении хвоста у головастика или при окукливании гусениц насекомых. Все эти процессы деградации катализируют главным образом ферменты, содержащиеся в лизосомах. Лизосомные ферменты способны разрушить большинство, а возможно, и все компоненты клетки, и поэтому заключены в непроницаемую мембрану, ограничивающую их активность. Поглощение клетками веществ высокомолекулярной природы на первых этапах протекает при участии плазматической мембраны, а затем лизосом, которые осуществляют деструкцию перевариваемых материалов.
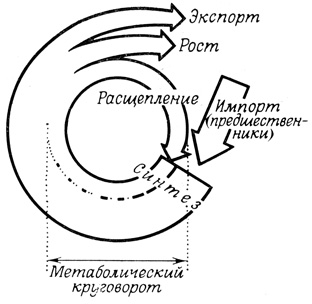
Рис. 19. Метаболический 'круговорот' веществ в клетке
Мы представили здесь крайне упрощенную картину функционирования клеток, описав лишь некоторые процессы, играющие ключевую роль во внутриклеточном хозяйстве. Значение этих процессов, характерных для большинства клеток, однако, сильно варьирует в зависимости от того, к какому типу принадлежит та или иная клетка (особенно это относится к клеткам сложных организмов). Внутренняя система клеток различного типа характеризуется собственным многообразием реакций. Это и есть тот путь, по которому осуществляется дифференцировка, в результате которой отдельные клеточные комплексы начинают выполнять специализированную роль в экономике целого организма (рис. 20).
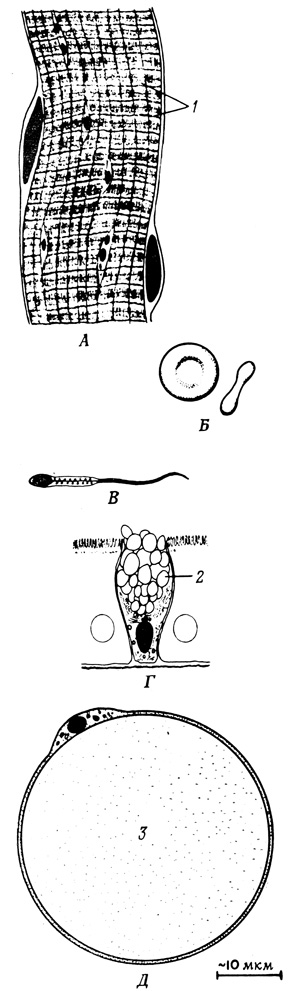
Рис. 20. Некоторые специализированные клетки высших животных. А - скелетная мышца. Б - эритроцит млекопитающих. В - сперматозоид. Г - бокаловидная клетка. Д - жировая клетка. 1 - мышечные нити; 2 - слизь; 3 - жир
Подобная специализация клеток проявляется часто в степени пролиферации мембраны, а также во взаимоотношениях между различными мембранными системами внутри клетки. Некоторые примеры специализации плазматической мембраны были уже отмечены выше. Вопросы, касающиеся митохондриальных мембран и высокоэнергетических процессов, протекающих в митохондриях, обсуждаются позже (гл. 5). Активность многих клеток сопряжена с синтезом и выделением во внешнюю среду макромолекул, используемых затем для различных внеклеточных целей. Так, связь между клетками осуществляется при помощи гормонов; в сборке внеклеточных структур участвуют предшественники клеточных стенок и коллаген; выделяемые во внеклеточное пространство муцины используются в качестве смазки; транспорт железа в тканях осуществляет трансферрин; узнавание и обезвреживание инородного материала - иммуноглобулины; наконец, выделяемые во внеклеточную среду ферменты и их предшественники используются для расщепления биологического материала. В процессах секреции принимают участие различные внутриклеточные органеллы, однако особая роль здесь принадлежит комплексу Гольджи.
О полярной (векторной) организации различных клеток мы уже упоминали. Подобная морфологическая асимметрия, при которой отдельные части клеток приспособлены для выполнения различных функций, наиболее выражена в эпителиальных клетках. Эти клетки граничат с двумя (или более) разделенными и физиологически обособленными средами. В таких случаях молекулы движутся через клетки только в одном направлении, причем движение это регулируется метаболизмом клетки и осуществляется благодаря специальным насосам, расположенным в клеточной мембране и контролирующим уровень различных растворимых веществ (гл. 2). В других случаях подвижные клетки, не имеющие морфологической полярности, могут, однако, ее приобрести под влиянием различных факторов окружающей среды. В каждом из описанных случаев полярности клеток она определяется и поддерживается работой различных механизмов, встроенных в клеточные мембраны.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'