2.7. Транспорт веществ при участии переносчиков, сопряженный с затратами метаболической энергии
Некоторые транспортные процессы, имеющие решающее значение для организма, протекают при участии специальных переносчиков и с затратами метаболической энергии. Это позволяет транспортировать вещества против химических или электрохимических градиентов. Такие процессы называют активным транспортом. Они удовлетворяют всем критериям, предложенным для идентификации процессов, протекающих при участии переносчиков. Таким образом, от других систем с облегченной диффузией активный транспорт отличается только тем, что он осуществляется против электрохимического градиента и связан с затратами метаболической энергии.
Возможность транспортировать вещества против концентрационных градиентов часто используется клетками для получения этих веществ из окружающей среды, где их концентрация чрезвычайно мала. В некоторых случаях это приводит к полному удалению потребляемых соединений из окружающей среды, например питательных веществ из сред, в которых культивируют бактерии, или из содержимого кишечника.
Градиенты - различных соединений, в особенности ионов, играют важную роль в метаболизме клеток. Создание и поддержание таких градиентов обусловлено процессами активного транспорта, которые помогают клеткам регулировать состав внутренней и внешней сред. Так, например, процессы активного транспорта, называемые часто натриевым насосом, поддерживают клетки в состоянии возбудимости. Этот насос обеспечивает низкое отношение натрий/калий в клетке, несмотря на то что во внеклеточной среде это отношение достаточно велико. Отношение градиентов Na+ и К+ равно в этом случае 15:1 (рис. 39). В других системах образуются градиенты значительно большего порядка. Так, клетки слизистой желудка с внутриклеточной концентрацией ионов Н+ около 10-7 М секретируют желудочный сок с концентрацией Н+ около 10-1 М. В данном случае концентрационный градиент ионов водорода составляет 106.
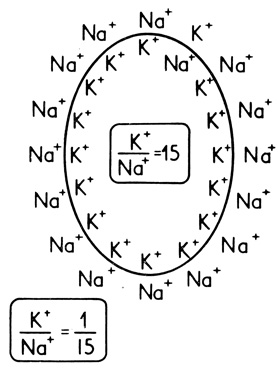
Рис. 39. Установившиеся градиенты ионов Na+ и К+, необходимые для осуществления некоторых функций клеток
Внутриклеточная концентрация Са2+, необходимая для сокращения скелетных мышц, составляет около 10-6 М. После сокращения концентрация Са2+ падает ниже 10-7 М, и мышца расслабляется. Это уменьшение внутриклеточной концентрации Са2+ осуществляется благодаря системам активного транспорта, локализованным в саркоплазматической сети. Когда мышца полностью приходит в расслабленное состояние, концентрация Са2+ в канальцах и цистернах саркоплазматической сети составляет, по-видимому, 10-3 М, а его концентрация в цитоплазме снижается до 10-7 М. Это соответствует концентрационному градиенту 104.
Сохранение таких концентрационных градиентов предполагает движение молекул и ионов в термодинамически невыгодном направлении, т. е. "в гору". Для такого движения требуются большие энергетические затраты, которые можно рассчитать по формуле
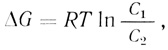
где ΔG - свободная энергия (работа), затрачиваемая на перенос 1 моля вещества против градиента концентрации (С1 - высокая концентрация; С2 - низкая концентрация). Величина концентрационного градиента определяет количество требуемой энергии. Последняя обеспечивается метаболическими процессами клетки, и поэтому вмешательство в энергетический обмен клетки приводит к ингибированию активного транспорта. Так, например, F- и иодацетат действуют на гликолиз; малонат и фторацетат - на цикл трикарбоновых кислот; CN-, N-3 - на дыхательную цепь; олигомицин и 2,4-динитрофенол - на окислительное фосфорилирование; аноксия - на дыхательную цепь, и наконец, охлаждение замедляет ферментативные реакции. Подобная взаимосвязь процессов может быть использована в качестве критерия, позволяющего судить об активном транспорте какого-либо иона или молекулы. Необходимо, однако, помнить, что эффективность данного ингибитора энергетического метаболизма зависит от характерного для данного типа клеток метаболизма, например в эритроцитах активный транспорт будут подавлять только ингибиторы гликолиза.
Необходимость использования больших количеств энергии для поддержания активного транспорта предъявляет высокие требования к интенсивности метаболизма клетки. В отдельных случаях на активный транспорт может затрачиваться почти вся энергия, вырабатываемая в клетке. У отдыхающего человека около 30-40% всей энергии метаболизма расходуется на активный транспорт. Уровень потребления кислорода является, по-видимому, наилучшим показателем интенсивности метаболизма в большинстве тканей, и важно отметить, что интенсивность потребления кислорода во многих тканях, в которых осуществляется активный транспорт, достаточно высока.
Хотя все без исключения клетки поддерживают ионные градиенты, нервная ткань занимает здесь особое место. В условиях часто повторяющейся деполяризации и сопровождающей ее утечки ионов нервные клетки должны постоянно восстанавливать ионные градиенты (гл. 3). Вес мозга человека составляет всего 1/50 веса тела, и тем не менее мозг отдыхающего человека потребляет около 1/5 всего количества кислорода, метаболизируемого организмом.
Почки человека продуцируют в день от 100 до 200 л клубочкового фильтрата, однако мочи выделяется всего лишь 1-2 л. Большая часть солей и практически все аминокислоты и сахара реадсорбируются из фильтрата. Физиологически активное вещество уабаин, ингибирующее главным образом активный транспорт натрия, на 70- 80% подавляет потребление кислорода почечной тканью. Из этого факта следует, что в почках на активный транспорт затрачивается основная часть метаболической энергии.
Некоторые процессы активного транспорта, например движение Na+, К+ и Са2+ через различные мембраны, а также трансмембранный перенос Н+ в митохондриях и бактериях могут быть непосредственно сопряжены с реакциями, продуцирующими энергию (гидролиз АТФ и окисление субстратов, рис. 40). В других случаях связь оказывается более сложной - в качестве движущей силы активного транспорта выступают ионные градиенты и электрические потенциалы, генерируемые первичными транспортными процессами, непосредственно сопряженными с расходом энергии.
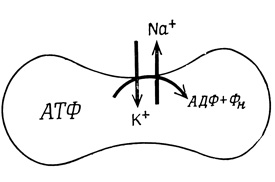
Рис. 40. Сопряженный с энергией транспорт ионов Na+ и К+
Микроинъекция АТФ или АТФ-синтезирующей системы в аксоны кальмара, отравленные цианидом или динитрофенолом, приводит к восстановлению натриевого насоса. Этот эффект можно наблюдать даже в "выпотрошенных" аксонах, из которых удалена цитоплазма. Роль АТФ в восстановлении натриевого насоса весьма специфична: многочисленные высокоэнергетические соединения оказались в этой системе абсолютно неэффективными. Замкнутые тени эритроцитов способны поддерживать натриевые и калиевые градиенты только в том случае, если в состав их внутренней среды входит АТФ. Транспорт натрия и калия сопровождается гидролизом АТФ, причем на каждую гидролизующуюся молекулу АТФ транспортируется приблизительно три иона натрия и два иона калия.
Первое наблюдение, указывающее на то, что транспорт ионов через мембрану может быть сопряжен с АТФазной реакцией, было сделано при изучении натриевого насоса в нерве краба. Мембранные фрагменты нерва краба не обладали заметной Мg2+-АТФазной активностью в присутствии ионов натрия или калия, однако совместное присутствие этих ионов приводило к значительному увеличению АТФазной активности. Последняя ингибировалась уабаином (рис. 41). Такие АТФазы, стимулируемые натрием и калием, были обнаружены в различных тканях. Активность этих ферментов и их кинетические свойства неразрывно связаны с активностью натриевых насосов клетки (табл. 1).
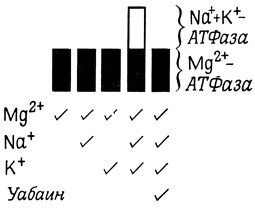
Рис. 41. Некоторые свойства (Na+ + K+) - зависимой АТФазы и Mg+ - зависимой АТФазы

Во многих тканях энергия, необходимая для транспорта различных метаболитов, может быть получена за счет ионных градиентов, генерируемых (Na++K+)-АТФазой. Накопление сахаров и аминокислот в кишечнике (рис. 42) и почках зависит, например, от присутствия Na+ во внешней среде, причем транспорт этих веществ ингибируется уабаином. Подобные эффекты объясняются совместным транспортом через мембрану Na+ и проникающей молекулы в виде тройного комплекса с переносчиком. Движущей силой такого транспорта (который протекает "в гору" для сахаров и аминокислот) служит градиент ионов натрия в условиях низкой концентрации натрия в клетке и при активно работающей (Na++К+)-АТФазе, способной поддерживать ионный градиент.
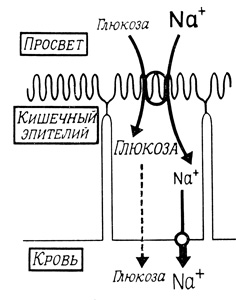
Рис. 42. Накопление глюкозы в клетках кишечного эпителия, зависящее от присутствия ионов Na+
Сейчас известно, что АТФазная активность связана также и с другими транспортными системами. Са2+-зависимая АТФаза определяет активность кальциевых насосов в саркоплазматической сети, мембране эритроцитов и в щеточной каемке кишечного эпителия. HCO-3-зависимая АТФазная Н+-секреция слизистой желудка связана с присутствием HCO-3-зависимой АТФазы.
Накопление Са2+ в митохондриях происходит либо за счет энергии АТФ, либо за счет транспорта электронов от дыхательных субстратов к кислороду по дыхательной цепи. Энергия запасается в форме высокоэнергетических промежуточных продуктов окислительного фосфорилирования (химическая гипотеза) (рис. 43) или при образовании протонного градиента, обусловленного векторным расположением дыхательных переносчиков (химио-осмотическая гипотеза) (гл. 5). Трансмембранные разности потенциалов, генерируемые некоторыми дегидрогеназами, могут также обеспечивать активный транспорт аминокислот в клетки E. coli.
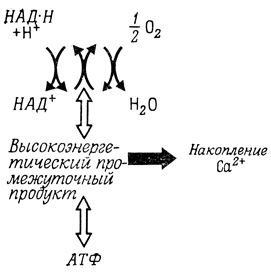
Рис. 43. Накопление ионов Са2+ в митохондриях
Механизм энергетического обеспечения транспорта ряда сахаров у некоторых бактерий включает фосфорилирование сахара на его пути в клетку. Фосфорилированные сахара накапливаются внутри клеток, причем донором фосфата служит фосфоенолпируват (ФЕП). Этот вид транспорта представляет собой транспорт определенной (гликозильной) группы через мембрану; однако он имеет много общих черт с энергозависимым транспортом, при котором химическая природа транспортируемых веществ не изменяется.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'