4.2. Синтез и хранение
Синтез полипептидных цепей, предназначенных для секреции, осуществляется на прикрепленных к мембранам рибосомах и имеет векторный характер. Синтезированные в результате этого процесса белки поступают в цистерны эндоплазматической сети. Такой механизм синтеза полипептидных цепей был выяснен (например, для синтеза амилазы в ацинарных клетках поджелудочной железы и для синтеза альбумина в печени) в опытах по выделению непроницаемых (для полипептидов) пузырьков шероховатой эндоплазматической сети из микросомных фракций, а также в экспериментах с пуромицином, который индуцирует отделение от рибосом недостроенных полипептидных цепей. Мембраны в выделенных пузырьках эндоплазматической сети ориентированы таким образом, что усеянная рибосомами поверхность смотрит наружу, а внутреннее замкнутое пространство соответствует внутреннему пространству цистерны эндоплазматической сети (рис. 80). Эти пузырьки были выделены из секретирующих тканей, кратковременно инкубированных с радиоактивными аминокислотами. Оказалось, что пузырьки несут связанные с рибосомами радиоактивные полипептидные цепи, представляющие собой белки, трансляция которых не успела закончиться за время гомогенизации. Если фрагменты шероховатой эндоплазматической сети обработать пуромицином, то большая часть таких полипептидных цепей окажется внутри микросомных пузырьков. Освобождаются эти полипептидные цепи только после разрушения мембраны пузырьков. Было высказано предположение, что в центре большой субчастицы рибосом и в мембране, с которой рибосомы связаны, имеется "дыра" и что растущие белковые цепи проходят через эти "дыры" в цистерны эндоплазматической сети. Одновременно связанная с мембранами ферментная система синтезирует дисульфидные связи в новообразованной молекуле белка (рис. 81).
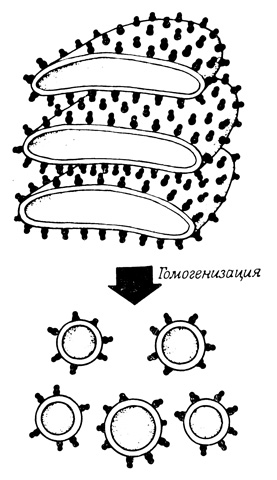
Рис. 80. Усеянные рибосомами мембранные пузырьки, которые образуются при гомогенизации шероховатой эндоплазматической сети
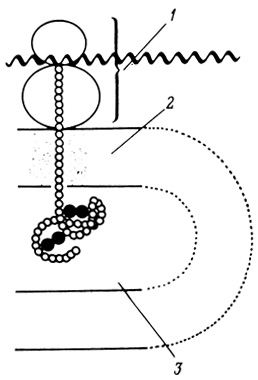
Рис. 81. Синтез полипептидной цепи. 1 - рибосома; 2 - мембрана; 3 - цистерна шероховатой эндоплазматической сети
Продукты, синтезированные на этой стадии, представляют собой "голые" белки, или полипептиды. Однако очень немногие клетки секретируют такие неизмененные белки, в большинстве же случаев эта исходная смесь "голых" полипептидов, прежде чем покинуть клетку, претерпевает различные модификации.
Первый и наиболее распространенный вид модификации - это добавление к полипептидной цепи различного числа боковых олигосахаридных цепей. Существует несколько типов олигосахаридных цепей, связанных с пептидной цепью через остатки аспарагина, треонина или серина. В результате присоединения большого числа углеводных цепей могут образоваться высокомолекулярные гликопептиды, состоящие главным образом из углеводов (например, муцины); если же число присоединившихся углеводных цепей невелико, то конечный продукт секреции изменяется очень незначительно. Последующая модификация углеводной части молекулы заключается часто в присоединении сульфатных групп, как это происходит, в частности, при образовании муцинов. Присоединение углеводных цепей к полипептидам осуществляют высокоспецифичные гликозилирующие ферменты, содержащиеся в мембранах эндоплазматической сети и главным образом комплекса Гольджи. Эти ферменты обращены, по-видимому, внутрь цистерн эндоплазматической сети и ламелл Гольджи. Внутреннее пространство указанных мембранных образований не сообщается с цитоплазмой, и поэтому не ясно, каким образом фосфорилированные сахара, необходимые для синтеза углеводной цепи, попадают в эти пузырьки. Вероятно, промежуточными продуктами синтеза полисахаридов здесь являются сахара, связанные с полипренол-фосфатами, как это имеет место при синтезе внеклеточных полисахаридов у микроорганизмов и растений.
Полипептиды, предназначенные для секреции, часто могут синтезироваться в такой форме, которая не обладает биологической активностью конечного продукта. Примером может служить инсулин, синтезируемый β-клетками островков Лангерганса поджелудочной железы (рис. 82). Продукт, который синтезируется эндоплазматической сетью, представляет собой проинсулин, состоящий из единичной полипептидной цепи и не проявляющий биологической активности. Биологически активным инсулин становится только после протеолитического удаления большого фрагмента (так называемого С-пептида) из центральной части полипептидной цепи. В результате расщепления полипептидной цепи, протекающего в комплексе Гольджи или в областях клетки, прилегающих к нему, образуется активный инсулин, состоящий из двух полипептидных цепей (цепи А и В), связанных между собой двумя дисульфидными связями. Некоторые другие полипептидные гормоны, такие, как окситоцин и вазопрессин (гормоны задней доли гипофиза), также, вероятно, образуются в результате протеолитического расщепления высокомолекулярных предшественников. Иногда полипептидные гормоны сразу синтезируются в активной форме; к числу таких гормонов относится меланоцитстимулирующий гормон.
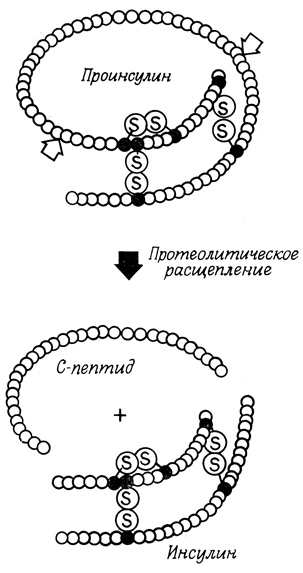
Рис. 82. Синтез инсулина в β-клетках островков Лангерганса поджелудочной железы
В клетках, секретирующих главным образом малые молекулы, такие, как нейромедиаторы или гистамин, продукты, которые накапливаются в пузырьках и высвобождаются в результате специфического воздействия на клетку, представляют собой смесь функционально активных малых молекул, специфических белков и сильно заряженных компонентов, например сульфатированных мукополисахаридов или смесь нуклеозидтрифосфатов. Такие дополнительные компоненты образуют, по-видимому, нерастворимые в воде комплексы с низкомолекулярными секретируемыми продуктами. Это позволяет клеткам накапливать большие количества секрета и не создает дополнительных проблем, связанных с поддержанием осмотического давления. Выше уже отмечалось, что высокомолекулярные компоненты секрета синтезируются, вероятно, в эндоплазматической сети и в комплексе Гольджи, однако происхождение физиологически активных низкомолекулярных соединений пока еще не выяснено. Например, в нервной системе запасающие гранулы синтезируются (и, вероятно, заполняются медиаторами нервного возбуждения) в теле нейрона. Затем они транспортируются по аксону к синапсам и далее высвобождаются из клетки в ответ на потенциал действия (рис. 83). Имеются данные (по крайней мере это известно для норадреналина), что выделившийся нейромедиатор может реадсорбироваться пресинаптическими окончаниями, поглощаться запасающими гранулами и снова высвобождаться наружу в ответ на стимул. В нервных окончаниях имеются также свои ферментные системы, осуществляющие синтез нейромедиаторов в дополнение к тем, которые поступают вместе с гранулами.
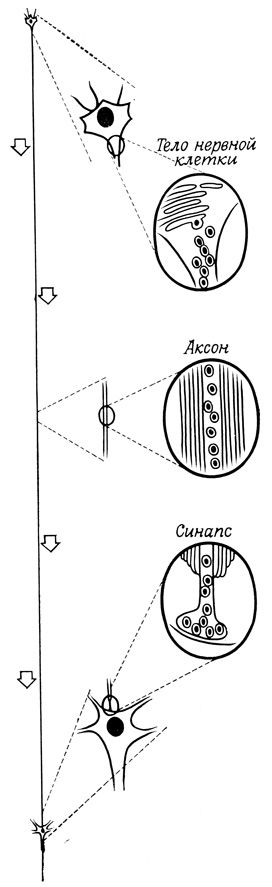
Рис. 83. Синтез и высвобождение нейромедиаторов
Некоторые белковые секреты находятся в гранулах в виде комплексов с ионами металлов (инсулин с Zn2+, амилаза околоушной слюнной железы с Са2+). Это, вероятно, облегчает создание высоких концентраций белков в запасающих гранулах.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'