4.1. Общая характеристика экзоцитоза
Лишь в небольшом числе случаев клетки секретируют низкомолекулярные соединения, несущие заряд, например ацетилхолин, катехоламины и гистамин; значительно чаще клетки секретируют макромолекулярные соединения, например белки плазмы и липопротеиды, переваривающие ферменты и их зимогены, полипептидные гормоны, компоненты клеточных стенок и муцины. Казалось бы, молекулы ни одного из этих веществ не могут свободно проходить через плазматическую мембрану. Высвобождение как малых, так и больших молекул из секретирующей клетки часто происходит в ответ на высокоспецифический внешний стимул, и это высвобождение обычно не сопровождается сколько-нибудь заметными потерями клеткой растворимых цитоплазматических компонентов, таких, как гликолитические ферменты.
Любое объяснение секреторной активности клеток основано на предположении о том, что либо для каждого типа секретируемого продукта существуют специальные молекулярные механизмы транспорта, либо клетки как-то умеют отделять материалы, предназначенные на "экспорт", от материалов, необходимых для "домашнего обихода", и затем, когда это требуется, высвобождать предназначенные на "экспорт" материалы. Об универсальности этого последнего механизма свидетельствуют данные, полученные при исследовании различных тканей.
Этот механизм иногда с некоторыми незначительными изменениями применим для объяснения синтеза, хранения и секреции разнообразных веществ, выполняющих различные функции (табл. 3). Список приведенных в табл. 3 веществ далеко не полон и служит лишь иллюстрацией, свидетельствующей о разнообразии таких систем.

Клетки, специализированные для секреции, обычно характеризуются некоторыми общими чертами (рис. 78). У этих клеток, во-первых, хорошо развита эндоплазматическая сеть, причем большая часть рибосом связана с мембранами этой сети; во-вторых, хорошо развит комплекс Гольджи и, наконец, в-третьих, имеется большое число гранул в цитоплазме, имеющих выраженную мембрану и заполненных, как правило, электроноплотным материалом. Одна из фракций частиц, выделяемых из гомогенатов секретирующих тканей методами субклеточного фракционирования, представлена главным образом гранулами с интактной мембраной. В этой фракции содержится значительная часть предназначенного для секреции материала. Гранулы, обычно локализованные вблизи той области клеточной мембраны, через которую осуществляется секреция, обеспечивают хранение секретируемых продуктов в клетке в форме, недоступной для других цитоплазматических компонентов до тех пор, пока эти продукты не высвободятся во внеклеточное пространство.
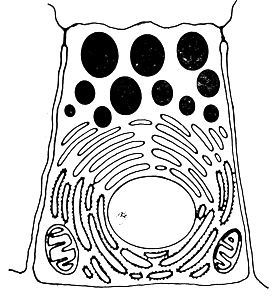
Рис. 78. Обобщенная схема клетки, специализированной для секреции
Общий механизм образования, хранения и высвобождения секретируемых продуктов для большого числа клеток (рис. 79) сейчас хорошо известен. Полипептидные компоненты всех секретируемых материалов синтезируются на рибосомах, прикрепленных к мембранам эндоплазматической сети. Далее эти компоненты транспортируются из цистерн эндоплазматической сети в цистерны комплекса Гольджи, где после необходимой химической модификации упаковываются в специальные пузырьки. К упакованным в пузырьках полипептидам добавляются низкомолекулярные компоненты, такие, как нейромедиаторы, нуклеозидтрифосфаты или ионы, необходимые для формирования белковых кристаллов; готовые гранулы либо непрерывно выбрасывают свое содержимое из клетки, либо остаются в цитоплазме до тех пор, пока соответствующий стимул не индуцирует высвобождения их содержимого во внеклеточное пространство путем экзоцитоза.
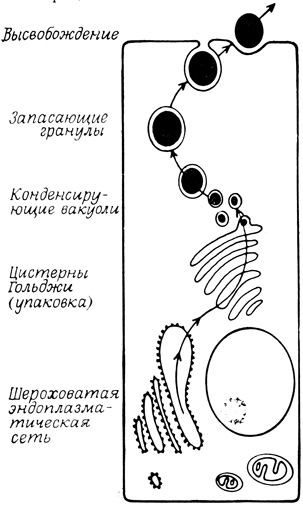
Рис. 79. Общая схема образования, упаковки и высвобождения секретируемых продуктов
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'