5.5. Структурные аспекты функционирования связанных с мембранами ферментов
Свойства ферментов, локализованных в мембранах, в различной степени зависят от окружения, которое создает для них интактная мембрана. Активность некоторых ферментов не изменяется даже в результате значительного упрощения молекулярной системы, в которой функционирует фермент, например при выделении фермента в виде индивидуального белка. Другие ферменты утрачивают свою активность даже при незначительных нарушениях мембранной структуры. Изучая условия, необходимые для поддержания, модификации и восстановления активности связанных с мембранами ферментов, исследователи одновременно получают важные данные о молекулярной организации мембраны.
Некоторые ферменты можно отделить от мембраны ("солюбилизировать") с помощью сравнительно мягкой процедуры. По-видимому, эти ферменты локализованы на поверхности мембраны. Большое число ферментов удается солюбилизировать только с помощью соединений, вызывающих значительные нарушения структуры мембран, например с помощью детергентов, органических растворителей и ферментов, гидролизующих фосфолипиды. Сохранение активности таких ферментов, вероятно, зависит от присутствия молекулярных агрегатов, в состав которых, помимо белков, входят еще липиды (или детергент).
Часто ферменты, отделенные от мембраны, не полностью проявляют биологические свойства, присущие им в составе мембраны. Это явление получило название аллотопии. Например, АТФаза (фактор F1) внутренней мембраны митохондрий утрачивает чувствительность к ингибитору олигомицину при отделении ее от мембраны. Чувствительность к ингибитору восстанавливается, если окружение АТФазы реконструировать с помощью соответствующих мембранных препаратов. Изменение свойств фермента при его отделении от мембраны в данном случае объясняется специфическим воздействием, которое оказывают на фактор F1 другие белки мембраны, причем воздействие этих белков на фактор F1 проявляется лишь в присутствии фосфолипидов мембран.
Липиды мембран являются важным фактором, обеспечивающим нормальное функционирование некоторых ферментов. Удаление этих липидов или изменение их свойств может отразиться на самых различных свойствах связанных с мембранами ферментов: от изменения кинетики ферментативной реакции или стабильности фермента до полной потери ферментативной активности. Поскольку потеря активности может быть обусловлена неспецифической денатурацией фермента, а не непосредственным участием липидов в реакции, всегда необходимо убедиться в том, что при добавлении липидов ферментативная активность восстанавливается.
Как правило, для восстановления ферментативной активности фосфолипиды более эффективны, чем нейтральные липиды. Во многих случаях восстановление ферментативной активности происходит лишь в присутствии больших количеств липида. Это справедливо, например, для ферментов дыхательной цепи митохондрий, цепи переноса электронов в микросомах, УДФ-глюкуронил-трансферазы и некоторых ферментов обмена липидов (фосфатидат - фосфогидролазы, стеароилдесатуразы, ацил-КоА-синтетазы, алкенил-ГФЦ - гидролазы). Количество липидов, необходимых для восстановления ферментативной активности, часто соответствует содержанию этого липида в исходной мембране (50-150 моль на 1 моль белка). В большинстве случаев ферменты не проявляют высокой специфичности к природе липидного материала, хотя каждая ферментная система может предпочитать некоторые индивидуальные липиды. Как правило, липиды необходимы для того, чтобы обеспечить соответствующее неполярное окружение для фермента - либо для поддержания его активной конформации, либо для облегчения доступа жирорастворимых субстратов к активному центру.
Некоторые ферментные системы требуют для своей работы наличия какого-то определенного липида, что свидетельствует о более специфическом взаимодействии белков и липидов в таких системах, причем, возможно, во взаимодействии участвуют не только гидрофобные части липидных молекул, но и соответствующие полярные группы. При этом часто эффективное воздействие на систему оказывают очень небольшие количества липида. В некоторых случаях восстановить ферментативную активность удается даже при добавлении 1-2 молекул липида на 1 молекулу белка (малатдегидрогеназа Mycobacterium avium и ГТФ-зависимая ацил-КоА - синтетаза из митохондрий печени крыс). Некоторые ферменты для проявления активности нуждаются в минорных липидах. По-видимому, присутствие этих липидов и объясняется наличием таких специфических взаимодействий. В клетках Escherichia coli обнаружен фермент, ФЕП-фосфотрансфераза (гл. 2), проявляющий свою активность только в присутствии фосфатидилглицерина, минорного липидного компонента этого микроорганизма. Если клетки E. coli или соответствующие фрагменты мембран обработать одной из разновидностей фосфолипазы D, которая специфически расщепляет фосфатидил-глицерин, то ФЕП-фосфотрансферазная активность исчезает; как только в результате биосинтеза фосфатидил-глицерин вновь появится в клетке, ферментативная активность восстанавливается.
Изменение липидного компонента мембраны, не приводящее к инактивации фермента, может, однако, изменить его свойства. Например, если на мембраны, содержащие аденилатциклазу, регуляция активности которой осуществляется гормонами, воздействовать ферментами, то активность фермента не изменится, однако чувствительность к гормонам полностью утрачивается. В некоторых случаях способность к ответу на гормоны удается восстановить при добавлении определенных липидов. Роль этих липидов сводится, возможно, к реассоциации белков регуляторной и каталитической субъединиц аденилатциклазного комплекса.
Некоторые процессы, протекающие на мембранах, характеризуются резким изменением кажущейся энергии активации при температурах, при которых наблюдаются также изменения физического состояния липидов мембран (фазовый переход). Температуру, при которой происходит изменение энергии активации процессов, протекающих на мембранах, в свою очередь можно изменить, изменяя состав остатков жирных кислот в мембране и тем самым изменяя ее физические свойства. Такие изменения в составе гидрофобного слоя мембраны можно индуцировать как у микроорганизмов, так и у животных, если на длительный срок изменить температуру содержания или уровень потребления различных жирных кислот (рис. 108). Иногда изменение энергии активации совпадает с фазовым переходом всей массы липидов мембраны. В этом случае изменение энергии активации ферментативного процесса отражает реакцию фермента на усредненное изменение состояния липидного окружения. В некоторых других случаях, однако, изменение энергии активации происходит при другой температуре, нежели фазовый переход липидов, но температура, при которой наблюдается изменение энергии активации, все еще зависит от липидного состава мембраны. Возможно, что в таких случаях на ферментативную активность оказывают влияние соседние с ферментом микрообласти липида, отличающиеся по своим физическим свойствам от липидной фазы остальной мембраны.
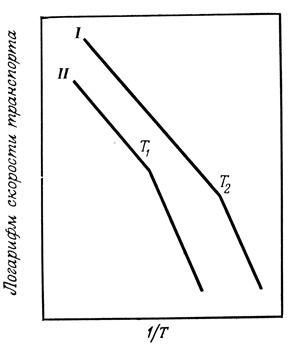
Рис. 108. Графики Аррениуса, характеризующие зависимость транспорта (β-глюкозидазы у бактерий, нуждающихся в ненасыщенных жирных кислотах, от температуры. I - рост на линолеате; II - рост на элаидате
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'