6.4. Жидкое состояние мембран
Мембранам приписывают "жидкую" структуру, поскольку многие компоненты свободно передвигаются в них. Плотная упаковка белковых молекул родопсина в мембранах фоторецепторов сетчатки, а также белка, осуществляющего транспорт Са2+ в мембранах саркоплазматической сети, не мешает этим белковым компонентам более или менее свободно вращаться вокруг своей оси, перпендикулярной плоскости мембраны, что было показано при изучении поляризации флуоресценции. Как уже отмечалось в гл. 3, имеются данные о том, что в клетках некоторых типов происходит миграция гликопротеидных молекул в плоскости мембраны. Эта миграция может осуществляться двумя способами, один из которых нуждается в притоке энергии, а другой - нет. Скорость вращательного движения и латеральной миграции всех этих молекул соответствует скорости свободной диффузии в вязкой среде. Миграция молекул рецептора ацетилхолина при образовании нервно-мышечных соединений служит еще одним примером латерального движения (см. ниже). Однако свободное движение присуще не всем мембранным белкам. Например, белковый компонент бляшек пурпурных мембран Halobacterium halobium образует геометрически регулярные плотноупакованные структуры, в которых ограничено вращательное движение. Сходные ограничения возникают, вероятно, в мембранах клеток мочевого пузыря и в областях межклеточных контактов. Подвижность белков ограничена также в тех случаях, когда для координации активности отдельных белковых компонентов мембраны требуются строго определенные пространственные взаимоотношения этих компонентов.
По некоторым данным, липидные компоненты мембран также обладают довольно большой свободой движения. Спектроскопические исследования показали, что углеводородные цепи липидных молекул обладают такой же свободой колебательных движений и гибкостью, какой обладают системы жидких углеводородов. Имеются, кроме того, данные о том, что диффузия липидов в плоскости мембраны протекает со скоростью, близкой к скорости диффузии соответствующих молекул в очищенных липидных системах. Лишь один тип движения молекул осуществляется, по-видимому, довольно редко - это перескок (или так называемый флип-флоп) молекулы с одной стороны мембраны на другую (рис. 123). Не только липиды, но и белки осуществляют такие перескоки не часто, поскольку для этого необходимы большие затраты энергии*.
* (Скорость флип-флопа, указанная на рисунке, относится к модельным фосфолипидным мембранам, построенным из одного только фосфатидилхолина. По некоторым данным, в биологических мембранах скорость флип-флопа может быть существенно выше. Флип-флоп фосфолипидов, по-видимому, ускоряется под влиянием таких веществ, как лизолецитин или окисленные липиды. Не исключено также, что молекулы некоторых белков, прошивающих мембрану насквозь, совершают вращательные движения, одновременно меняя свою конформацию, в результате чего их наружные сегменты попадают внутрь, и наоборот. - Прим. ред.)
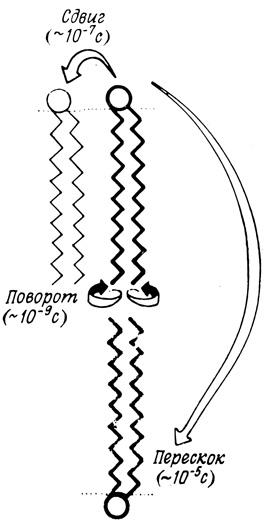
Рис. 123. Различные типы движений липидных молекул
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'