6.7. Включение новосинтезированных компонентов в мембрану
Попытки реконструировать мультиферментные системы из изолированных мембранных компонентов показали, что наилучших результатов можно добиться в том случае, если мембранные белки добавлять к сформированным заранее липидным структурам, содержащим липиды нужного типа. Включение различных белковых компонентов в эти структуры может осуществляться либо одновременно, либо последовательно. Кроме того, эксперименты, проведенные с бактериальными мутантами, не способными синтезировать фосфолипиды при отсутствии в культуральной среде глицерина, показали, что для включения в существующую мембрану новой транспортной системы не требуется увеличения мембраны за счет синтеза новых молекул фосфолипидов. Следует, однако, отметить, что функциональная активность транспортной системы у таких мутантов часто может быть снижена. При выращивании бактериальных мутантов, нуждающихся в жирных кислотах, на средах, содержащих те или иные жирные кислоты, можно индуцировать изменения в физических свойствах мембранных липидов. Для каждого типа клеток существует некая критическая температура, ниже которой белки транспортных систем не могут включаться в клеточные мембраны. Создается впечатление, что одним из условий включения белков в мембраны является жидкое состояние мембранных липидов. Добавление к охлажденным ниже критической температуры клеткам веществ, индуцирующих образование транспортной системы в клеточной мембране, вызывает синтез белков этой транспортной системы в клетке. Однако синтезированные белки не включаются в мембрану, и явление индуцированного транспорта, таким образом, не наблюдается. Транспортная система не образуется и при последующем повышении температуры.
Независимый механизм включения различных компонентов в мембрану хорошо согласуется с тем, что обновление этих компонентов в одной и той же мембране протекает с различными скоростями, а также с тем, что ново-синтезированные компоненты сначала равномерно распределены по всей мембране. Остается, однако, непонятным, каким образом в физически непрерывной мембране формируются отдельные области, резко отличающиеся от остальных содержанием определенных белков.
Подобная ситуация возникает, например, при размножении вирусов, имеющих оболочку. Синтез вирусных нуклеокапсидов сопровождается одновременным включением в плазматическую мембрану клетки-хозяина белков и гликопротеидов, которые впоследствии войдут в состав оболочки вируса. Эти белки и гликопротеиды формируют в мембране небольшие области, в которых не содержится обычных белковых компонентов плазматической мембраны клетки-хозяина. Когда нуклеокапсиды готовы, они "заселяют" области плазматической мембраны, сформированные из вирусных белков. Эти области мембраны и образуют в дальнейшем оболочку вируса. Готовые вирусы отпочковываются от плазматической мембраны и уходят во внеклеточное пространство, унося с собой часть мембранного материала клетки (рис. 126). Таким образом, вирусы приобретают оболочки, состоящие из вирусных белков и гликопротеидов, включенных в липидный матрикс плазматической мембраны клетки-хозяина.
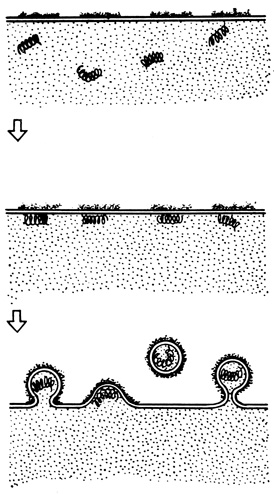
Рис. 126. Образование оболочки у некоторых форм вирусов
В процессе дифференцировки эмбриональной мышечной ткани изменяется распределение рецепторов ацетилхолина на поверхности клеток. Первоначально эти рецепторы равномерно распределены по поверхности клеток, однако в процессе дальнейшей дифференцировки клеток рецепторы образуют на мембране небольшие области, с которыми взаимодействуют растущие нервные аксоны, образуя нервно-мышечное соединение.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'