6.9. Регуляция
Характер мембран, а также скорость их обновления могут определяться исключительно скоростями синтеза их индивидуальных компонентов. Например, если в культуру добавить какое-то питательное вещество, то в бактериальных клетках развиваются соответствующие системы транспорта с участием переносчика, локализованного в мембране. Этот процесс включает стадию биосинтеза компонентов транспортной системы и их последующее внедрение в мембрану. С момента введения в среду питательного вещества скорость индуцированного транспорта в клетке возрастает, пока не достигнет определенного постоянного уровня. Этот уровень соответствует состоянию равновесия в клетке, при котором скорость синтеза равна скорости деградации.
Регуляция в клетках млекопитающих, осуществляемая в основном гормонами, имеет более сложный характер и более точно сбалансирована. Например, в клетках печени новорожденной крысы содержится морфологически развитая эндоплазматическая сеть, однако в мембранах этой сети отсутствуют функционально активные ферменты системы транспорта электронов. В течение первой недели жизни после рождения происходит развитие этой системы, причем мембраны не меняют своего вида (рис. 127). Включение различных компонентов системы транспорта электронов в мембрану происходит асинхронно. Таким образом, есть основания предполагать, что отдельные компоненты включаются в мембрану независимо друг от друга, а не в виде структурно организованных мультиферментных комплексов.
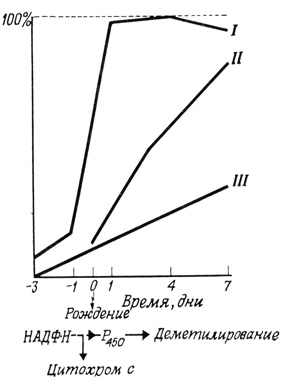
Рис. 127. Развитие системы транспорта электронов в клетках печени крысят. По оси ординат отложены относительные величины активности различных ферментных систем (в % от уровня активности взрослых животных). I - активность НАД⋅Н - цитохром-с-редуктазы; II - активность Р450; III - степень деметилирования
Регуляция такого же типа наблюдается и в ответ на аномальный стимул, например на воздействие фенобарбитала (гл. 5). В этом случае в течение нескольких дней происходит интенсивное развитие гладкой эндоплазматической сети, содержащей ферменты, способные катализировать расщепление фенобарбитала. Образование новых мембранных структур требует увеличения уровня фосфолипидов и холестерина. Удивительно, что накопление этих липидов в клетке происходит за счет уменьшения скорости деградации фосфолипидов и увеличения скорости синтеза холестерина. При этом скорость деградации холестерина не меняется, а скорость синтеза фосфолипидов если и меняется, то в сторону уменьшения. Увеличение содержания мембранных ферментов достигается в большинстве случаев путем одновременного возрастания скорости биосинтеза и уменьшения скорости деградации.
Скорость, с которой клетка отвечает на функциональные требования, предъявляемые к одной из ее мембранных структур, можно оценить по скорости изменений, происходящих с плазматической мембраной макрофага после фагоцитоза (рис. 128). В процессах эндоцитоза (или пиноцитоза) значительная часть плазматической мембраны перемещается внутрь клетки, формируя мембраны фагоцитозных вакуолей. Эти клетки, характеризующиеся обычно большой подвижностью поверхности и высоким отношением поверхность/объем, становятся округлыми и утрачивают способность к фагоцитозу. В течение последующих нескольких часов происходит понижение уровня 5'-нуклеотидазы в проглоченной мембране (время полужизни ≈2 ч), однако концентрация холестерина и фосфолипидов остается постоянной. Через 4-10 ч после фагоцитоза развивается новая плазматическая мембрана (доказательством этому служит параллельное увеличение уровня 5′-нуклеотидазы, холестерина и фосфолипидов), поверхность клетки увеличивается и восстанавливается ее способность к фагоцитозу и пиноцитозу. Степень интеграции функции, присущая этому процессу, становится особенно заметной при сравнении обычных клеток и клеток, выращенных в среде, не содержащей холестерина (макрофаги не способны синтезировать холестерин из ацетата). В таких лишенных холестерина клетках восстановления указанных выше функций не наблюдалось.
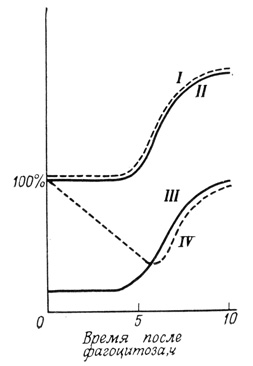
Рис. 128. Изменения, происходящие в макрофагах после фагоцитоза. По оси ординат отложены относительные величины (в % от нормального уровня), характеризующие состояния макрофага и состав его мембраны. I - содержание фосфолипидов; II - содержание холестерина; III - способность к фагоцитозу; IV - содержание 5′-нуклеотидазы
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'