Развитие межклеточных контактов в фило- и онтогенезе (Л. В. Гербильский)
Системообразующая функция межклеточных контактов в филогенезе животных (Л. В. Гербильский)
Исследователями школы Л. А. Орбели установлено, что патологические изменения, происходящие в системах организма, сопровождаются нарушением взаимодействий элементов этих систем. Разрушаются взаимоотношения элементов, характерные для более поздних ступеней эволюционного развития, и устанавливаются новые, патологические взаимодействия, повторяющие более древние этапы филогенеза.
Сказанное делает понятным интерес исследователей к сравнительному изучению контактных взаимоотношений клеток животных, находящихся на различных ступенях эволюционного развития (U. Welsch, W. Buchheim, 1978). По мнению А. Г. Маленкова, Г. А. Чуича (1979), одна из важных задач контактологии - выяснение характера и роли изменений МК в эволюционном процессе. Решение этой задачи может способствовать пониманию роли контактных взаимодействий при патологических состояниях.
U. Welsch, W. Buchheim (1978) считают, что имеющиеся в настоящее время данные не позволяют выявить основные линии филогенетического развития контактов. Структура МК до сих пор изучена у небольшого числа представителей животного мира, однако развиваемое нами представление о МК как о системообразующих элементах тканевых систем (В. И. Архипенко и др., 1979) дает возможность систематизировать накопленные факты, основываясь на единой теоретической концепции, и обнаружить некоторые закономерности эволюции МК. Исходя из этого, Л. В. Гербильский (1980) проанализировал эволюционную динамику МК и установил основную закономерность эволюции данных структур: повышение системообразующей функции МК в процессе эволюции. На основании этого можно сформулировать основные задачи сравнительной контактологии.
Первая задача сравнительной контактологии - выяснение закономерностей первичных контактных взаимодействий клеток, благодаря которым стало возможным появление многоклеточных животных. У современных одноклеточных (в частности, у жгутиковых) имеются специализированные структуры клеточной поверхности, которые могли быть использованы при формировании контактов между клетками примитивных многоклеточных. Такие структуры у жгутиковых, обеспечивающие стабилизацию жгутика при движении, находятся между плазмолемма- ми жгутика и клеточного тела и соответствуют десмосомам (A. S. Brooks, 1978). Эти десмосомоподобные соединения расположены несколькими рядами по 3-5 элементов. Два противостоящих подмембранных уплотнения каждого элемента имеют диаметр 15-30 нм и локализуются с интервалом 50-100 нм. Межмембранное пространство данных элементов шириной 10-40 нм заполнено аморфным материалом, а у некоторых жгутиковых выявлена центральная электронноплотная линия.
Элементы типа гемидесмосом образуются у жгутиковых для обеспечения взаимодействия с различными структурами, в частности у паразитических форм,- с клетками и хозяина. С помощью десмосомоподобных структур клетки различных жгутиковых могут взаимодействовать друг с другом, образуя многоклеточные комплексы (H. Ritter и др., 1978). Более сложно организованные соединения располагаются между клетками сизигия грегарин. Эти соединения напоминают септированные и участвуют в адгезии двух клеток грегарины (Е. С. Снигиревская и др., 1977; Е. С. Снигиревская, Я. Ю. Комиссарчик, 1980). Приведенные данные позволяют предполагать, что филогенетйчески наиболее ранней функцией МК явилось обеспечение механических взаимодействий между клетками. Однако систематическое исследование МК У наиболее низко организованных многоклеточных животных пока не проведено. По-видимому, изучение конъюгации у простейших может дать интересную информацию о становлении межклеточных взаимодействий у многоклеточных животных (P. Luporini и др., 1979).?
Сравнительный анализ контактных взаимодействий, изученных на различных моделях, позволит внести важный вклад в решение одной из основных проблем биологии - проблемы перехода от одноклеточных организмов к многоклеточным.
Вторая задача, стоящая перед сравнительной контактологией,- установление морфологических закономерностей повышения системообразующей функции МК. Полученные в последние годы данные позволяют условно выделить три этапа эволюции контактов: возникновения межклеточных соединений, их специализации и образования специализированных комплексов (рис. 30).
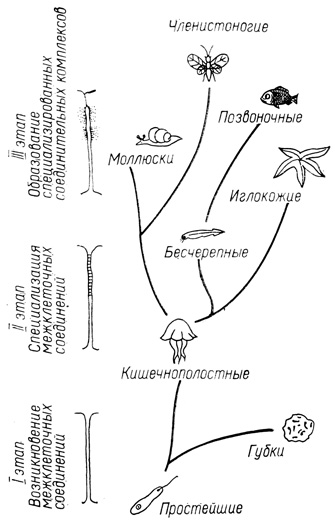
Рис. 30. Основные этапы эволюции системообразующей функции МК
Первый этап в настоящее время представлен, вероятно, наиболее примитивными многоклеточными-губками. Из структурных элементов МК губок следует назвать прежде всего неспециализированные простые соединения с параллельным расположением плазмолемм, между которыми локализуется мукополисахаридная прослойка, окрашивающаяся рутением красным. Наряду с простыми у известковых губок имеются соединения, которые можно отнести к классу септосодержащих. Они располагаются между склеробластами, участвующими в формировании одной спикулы. У губок данные соединения обеспечивают взаимодействие нескольких склеробластов, что приводит к образованию трех- или четырехлучевой спикулы. Хотя между клетками губок существует электрическая связь, у них пока не найдены щелевые соединения. Истинные плотные соединения и десмосомы у губок также, по-видимому, отсутствуют.
Таким образом, системообразующая функция контактов у губок развита незначительно. Контактные взаимодействия обеспечиваются главным образом путем формирования временных или постоянных простых соединений между плазмолеммами. Эти соединения не фиксируют клетки жестко, а позволяют им смещаться относительно друг друга. Адгезия клеток незначительна, поэтому организм губок легко разделить на отдельные клетки.
Агрегация клеток губок осуществляется с помощью АФ, структура которых в настоящее время достаточно полно изучена (E. G. W. Miller и др., 1979). Уже через 1-2 ч между агрегирующими клетками губок можно наблюдать формирование различных межклеточных соединений, напоминающих десмосомальные и другие специализированные соединения высших многоклеточных животных (S. Kartha, S. Mookerjee, 1979). Изолирующая функция МК у губок отсутствует - у них не образуется изолированная внутренняя среда. Коммуникация между клетками губок, по-видимому, незначительна.
Второй этап эволюции контактных взаимодействий связан со специализацией клеточных мембран, межмембранных и подмембранных компонентов в трех направлениях: увеличения адгезии, коммуникации и изоляции (см. схему). Простые (неспециализированные) соединения в определенной степени обеспечивают все три аспекта системообразующей функции, однако ее совершенствование потребовало возникновения специализированных соединений (адгезивных, коммуникативных, изолирующих).
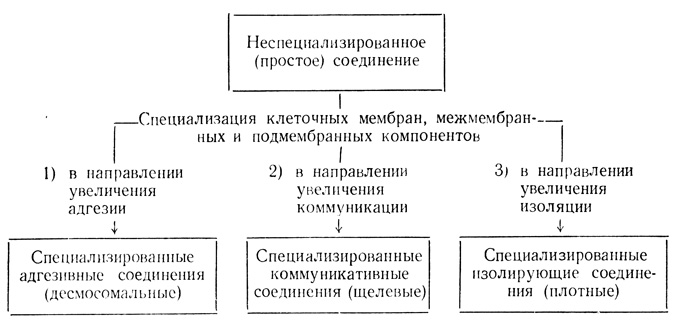
Гипотетическая схема развития системообразующей функции МК в филогенезе животных
Уже у кишечнополостных животных хорошо развиты все три типа соединений. Наиболее изучены контактные взаимодействия клеток гидры (R. L. Wood, 1977; Filshie, Flower, 1977).
Апикальные участки плазмолемм клеток в экто- и энтодерме связаны между собой хорошо выраженными септированными соединениями длиной до 4 мкм, состоящими из многочисленных параллельных септ. Функция септированных соединений - изоляция, образование барьера, ограничивающего межмембранное пространство контакта. Маркерное вещество (пероксидаза хрена) при добавлении в среду не проникает в межмембранное пространство между клетками. Однако это становится возможным при предварительной обработке гидр проназой, приводящей к расщеплению септ без нарушения целостности плазмолемм. Септированные соединения способны быстро восстанавливаться при регенерации гидр после дезинтеграции. Процесс восстановления септированных соединений при регенерации гидр подробно описан. Вполне вероятно и участие этих соединений в адгезии (D. Whalee и др., 1978; R. L. Wood, A. M. Kuda, 1978; M. Brim и др., 1980; R. L. Wood, 1980).
Чрезвычайно развиты у гидр щелевые соединения, соединяющие миоэпителиоциты экто- и энтодермы. По-видимому, эти специализированные коммуникативные соединения впервые появляются у кишечнополостных. Протяженность щелевых соединений достигает 4 мкм, они располагаются группами, и между двумя соседними клетками может быть до 5 таких соединений, плотно прилежащих одно к другому. Применение метода криоскалывания позволило установить, что щелевые соединения гидр состоят из шестиугольных частиц, разделенных промежутками. В связи с небольшим числом нейроцитов у кишечнополостных щелевые соединения могут обеспечивать синхронизацию расслабления и сокращения отдельных клеток (R. J. Wakeford, 1979).
Щелевые соединения обнаружены также между тонкими отростками миоэпителиоцитов (Д. В. Гуляев, 1976). Очевидно, именно через эти соединения красители могут проникать из одних миоэпителиоцитов в другие. Иной класс соединений - десмосомальные - формируется между толстыми отростками миоэпителиоцитов. Эти соединения выполняют механическую, адгезивную функцию. Механическую функцию осуществляют также десмосомы, фиксирующие нематоциты гидромедуз. К десмосомам прикрепляются многочисленные пучки филаментов нематоцитов. Высокая степень адгезии в данном месте препятствует отрыву нематоцита вместе с захваченной им жертвой. Десмосомы, септированные и щелевые соединения связывают клетки кораллов и антомедуз (M. G. King, A. N. Spencer, 1979). У ктенофор также имеются септированные, щелевые и десмосомальные соединения.
Межклеточные взаимодействия свободноживущих представителей плоских червей изучены мало (V. Storch, U. Welsch, 1977). У планарии обнаружены септированные, щелевые и десмосомальные соединения (D. С. Quick, R. G. Johnson, 1977). Установлено, что септированные соединения выполняют, подобно плотным соединениям позвоночных, изолирующую функцию. Лучше исследованы МК паразитических плоских червей. Между клетками этих животных также встречаются септированные и щелевые соединения. У представителей данного типа много-клеточных животных, по-видимому, впервые появляются гемидесмосомы. Предполагают, что это связано с возникновением системообразующих элементов органного уровня - базальных мембран, координирующих пространственное расположение эпителиоцитов и клеток подлежащей ткани (В. И. Архипенко, Л. В. Гербильский, 1978). К гемидесмосомам прикрепляются филаменты, которые могут обеспечивать ориентацию органелл эпителиоцитов по отношению к базальной мембране. С другой стороны, базальная мембрана служит местом прикрепления коллагеновых волокон или мышечных клеток, которые фиксируются также с помощью гемидесмосом.
У трематод между клетками кишечника и паренхимы выявлены многочисленные соединения, в которых прилежащие клеточные поверхности располагаются параллельно на расстоянии 20 нм. Возможно, по этим соединениям у данных животных, не имеющих сосудов, осуществляется транспорт метаболитов. У киноринхов эпителиоциты эпидермиса связаны между собой септированными и десмосомальными соединениями; десмосомы выявлены также у немертин (М. В. Атаманова, 1980).
У представителей типа кольчатых червей прикрепление эпителиоцитов к базальной мембране также осуществляется с по-мощью гемидесмосомы. Десмосомальные соединения найдены у полихет и олигохет (W. Binge и др., 1979), причем у некоторых кольчатых червей обнаружены необычные десмосомы, отличающиеся значительной шириной межмембранного пространства. Септированные соединения выявлены у олигохет, приапулид и коловраток (D. G. Baskin, 1976). Недавно у кольчатых червей обнаружены щелевые соединения (A. Bilbaut, 1980).
Таким образом, у кишечнополостных, а затем у низших представителей двустороннесимметричных животных - плоских, круглых и кольчатых червей - наблюдается процесс специализации межклеточных соединений, приводящий к образованию специализированных элементов контакта. Щелевые соединения особенно характерны для кишечнополостных, хотя у червей эти элементы также участвуют в функциональном объединении эпителиоцитов (A. Bilbaut, 1980).
Третий этап, характеризующий взаимодействие клеток высших представителей первичноротых и вторичноротых животных, заключается в формировании принципиально новых структур - соединительных комплексов, состоящих из нескольких специализированных соединений. Не исключено, что такие комплексы образуются и на более ранних этапах эволюции, но среди первичноротых животных они особенно характерны для моллюсков и членистоногих, а среди вторичноротых - для иглокожих. Главную роль в комплексах играют изолирующие (плотные и септированные) соединения. В связи с развитием у высших животных сосудистой и скелетной систем коммуникативные и адгезивные соединения в определенной степени теряют ведущее значение в организации контакта. Соединительные комплексы моллюсков расположены между апикальными частями эпителиоцитов и включают септированные соединения (J. M. Arnold и др., 1978). Своеобразные комплексы обнаружены между эпителиоцитами жабр двустворчатых моллюсков. У этих животных септированные соединения чередуются со щелевыми так, что между соседними септами плазмолеммы клеток сближаются до 2-4 нм, образуя локальные щелевые соединения.
Другой тип соединительных комплексов расположен между эпителиоцитами мантии. Этот комплекс состоит из десмосомы и септированного соединения. Базальнее находится простое соединение, в котором имеются локальные расширения до 1 мкм. В этих расширениях наблюдаются электронноплотные гранулы диаметром 10-20 нм, в составе которых с помощью электронной цитохимии обнаружено большое количество Ca. Предполагают, что описанная структура контакта предназначена для транспорта Ca на поверхность эпителия, что необходимо для построения раковины. Содержащие Ca гранулы образуются в субэпителиальной соединительной ткани, затем поднимаются по контакту до уровня септированного соединения, поступают в цитоплазму и через микроворсинки выводятся в мантийную полость.
Значительное число исследований посвящено изучению межклеточных взаимодействий у членистоногих. Подробно исследована структура МК эпителиоцитов печеночного выроста ракообразных (M. V. Conzales, 1976). Эти контакты образованы септированными и щелевыми соединениями. Септированные соединения у дафний опоясывают апикальную треть эпителиоцита и имеют протяженность 10 мкм. Межмембранное пространство шириной 18 нм заполнено материалом с повышенной электронной плотностью. Септы располагаются на расстоянии 3,5 нм друг от друга. Ниже септированных соединений плазмолеммы контактирующих клеток разделены межмембранным пространством 15-50 нм. Здесь локализуются отдельные щелевые соединения протяженностью около 0,5 мкм.
Иначе построены МК эпителиоцитов гепатопанкреаса Oniscus ascellus и Ascellus interrnedius. Латеральные поверхности апикальных частей этих клеток связаны соединительными комплексами, состоящими из протяженной зоны слипания и соединения, напоминающего плотное.
МК жабр креветок образованы тремя видами соединений: зонами слипания, септированными и напоминающими плотные. В зонах слипания, имеющих длину 0,1-0,2 мкм, межмембранное пространство расширяется до 25 нм и заполнено электрон- ноилотным содержимым. Прилежащая к этому соединению цитоплазма также уплотнена. Септированное соединение протяженностью 0,8 мкм и больше располагается базальнее. На поперечных срезах видны септы шириной 7 нм, отделенные друг от друга промежутком 8 нм. Межмембранное пространство имеет ширину 20 нм. На тангенциальных срезах это соединение имеет вид пчелиных сот. В различных участках контакта встречаются соединения, напоминающие плотные. Подобные соединения находятся также между эпителиальными и трабекулярными клетками. Десмосомальные и щелевые соединения между эпителиоцитами не обнаружены. Сходные соединительные комплексы, состоящие из области слипания и септированного соединения, выявлены также между эпителиоцитами протока максиллярной железы других рачков.
Контакт между клетками эпидермиса Limulus образован септированными соединениями и зонами слипания, а у Balanus - десмосомами и септированными соединениями. Десмосомы связывают также базальные поверхности сухожильных эпителиоцитов с миоцитами. Однако эти элементы отличаются от классических десмосом значительной шириной (25-70 нм) межмембранного пространства, заполненного материалом с незначительной электронной плотностью. С кутикулой эпителиоциты связаны посредством гемидесмосом. Подобным образом через эпидермис осуществляется связь между мышцами и кутикулой у других ракообразных и пауков. Следует отметить важную роль МК в транспорте ионов Са+2 у ракообразных, имеющих обызвествленный наружный скелет.
Своеобразие взаимоотношений эпителиоцитов шейных органов личинок некоторых рачков определяется функцией этих клеток - экскрецией ионов Na+. Эпителиоциты контактируют друг с другом только в апикальной области с помощью септированных соединений. Ниже контакт между клетками не наблюдается, а латеральные плазмолеммы образуют многочисленные складки, вдающиеся в цитоплазму. Аналогичную структуру имеет МК хлоридных клеток насекомых. Подобные складки плазмолеммы формируются в области МК между солевыми клетками рыб и клетками солевых желез водоплавающих птиц (C. V. Riddle, S. A. Ernst, 1979; C. Sardet и др., 1979; S. DunelErb, P. Laurent, 1980). Между клетками кишок различных представителей ракообразных обнаружены септированные и щелевые соединения, а также зоны слипания. Септированные соединения найдены и у морских пауков (C. R. Green, 1981).
Особенно хорошо изучены контактные взаимоотношения клеток у насекомых. В эпидермисе этих животных присутствуют септированные соединения и десмосомы. Одни авторы отмечают определенную закономерность в размещении этих элементов на контактной поверхности клеток, другие же отрицают ее (D. J. Beadle, M. W. J. Megaw, 1979; J. C. Andries, 1979).
На тангенциальных срезах видно, что, как правило, септированные соединения насекомых отличаются по структуре от подобных элементов контакта клеток кишечнополостных. На таких срезах видна сеть напоминающих пчелиные соты шестиугольных ячеек диаметром 10-13 нм, с плотными стенками и светлой центральной частью. В связи с этим данные соединения целесообразно называть ячеистыми. Ширина межмембранного пространства ячеистых соединений равна 14-30 нм (Л. В. Данилова и С. Н. Селезнева, 1967).
Эпителиальные клетки слюнных желез и некоторых других органов насекомых соединяются с помощью ячеистых соединений. В других случаях септированные соединения между клетками насекомых подобны соединениям кишечнополостных и на тангенциальных срезах представлены параллельными линиями. На сколах септированных соединений обнаруживаются внутри-мембранные частицы, расположение которых частично соответствует расположению септ. Поскольку под мембранами септированных соединений располагаются уплотнения цитоплазмы, можно предполагать, что внутримембранные частицы обеспечивают прикрепление септ к этим уплотнениям (C. Noirot-Timothee и др., 1978). Строение септированных соединений у различных представителей беспозвоночных вариабельно. Можно выделить не менее 8 разновидностей данных элементов. Одной из актуальных задач в области изучения этих соединений является создание их классификации с учетом данных, полученных с помощью криоскалывания (С. R. Green, 1978, 1981).
Септированные соединения наиболее характерны для МК эпителиоцитов пищевого канала насекомых (C. Noirot-Timothee и др., 1978; D. E. Bignell, 1980). Только у некоторых чешуекрылых септированные соединения между эпителиоцитами кишок не обнаружены. В кишках так же, как и в эпидермисе, встречаются щелевые соединения и десмосомы. В одних случаях эти элементы располагаются упорядоченно, образуя соединительные комплексы, в других - такой закономерности выявить не удается. С помощью метода криоскалывания обнаружены значительные отличия в структуре щелевых соединений насекомых и других членистоногих по сравнению со щелевыми соединениями иных типов. У последних внутримембранные частицы диаметром 7-8 нм находятся на внутренней поверхности скола в виде характерных упорядоченных полигональных скоплений (щелевое соединение типа Р). У членистоногих более крупные внутримембранные частицы диаметром 8-12 нм располагаются на наружной поверхности скола и не образуют полигональных скоплений - щелевое соединение типа Е, или инвертированное (N. E. Flower, 1977).
Десмосомы насекомых отличаются от десмосом других животных тем, что к этим соединениям прилежат микротрубочки, размещенные параллельно клеточной поверхности. В составе МК кишок некоторых насекомых выявлены два вида своеобразных соединений. Одно из них получило название непрерывного (R. Dallai, 1976) и отличается параллельностью плазмолемм соседних клеток, лежащих на растоянии 11 -15 нм. Межмембранное пространство заполнено аморфным электронноплотным содержимым гликопротеидной природы и пронизано тонкими поперечно расположенными фибриллами, которые соединяют между собой плотные гранулы, лежащие под плазмолеммами контактирующих клеток. При введении La в межмембранном пространстве непрерывного соединения выявлены поперечно расположенные септы толщиной 4-5 нм. Эти элементы обнаружены также между клетками сперматеки насекомых и эпителиоцитами ракообразных. Очевидно, эти соединения прочно связывают клетки, не позволяя погибающим эпителиоцитам отделяться во время линьки, и сохраняют целостность эпителиального пласта (A. Krzysztofowicz, S. Kuzyk, 1979).
Другое соединение, выявленное в кишках и различных отделах выделительной системы насекомых, получило название лестничного. В области его межмембранное пространство шириной 20 нм пронизано трубочками, расположенными поперечно на расстоянии 25 нм друг от друга. Возможно, лестничное соединение также выполняет функцию адгезии (С. Noirot-Timothee и др., 1979).
Важно выяснить, у каких животных впервые появляются плотные соединения. Долгое время считалось, что эти элементы МК формируются только у хордовых и отсутствуют у представителей иных типов (U. Welsch, W. Buchheim, 1978). Однако применение методики криоскалывания позволило установить, что в некоторых органах насекомых присутствуют плотные соединения, хотя они развиты в меньшей степени, чем у позвоночных животных (N. J. Lane, 1981).
На Р-поверхности сколов семенника шелкопряда обнаружены зигзагообразные ряды частиц, а на Е-поверхностях - комплементарные им ряды углублений. Плотные соединения создают эффективный барьер между гемолимфой и развивающимися половыми клетками шелкопряда (K. Toshimori и др., 1979). Подобные плотные соединения формируют барьер между гемолимфой и клетками мозга, а также сетчатки у мух (С. Shi, D. T. Carlson, 1980). Таким образом, у высших первичноротых животных плотные соединения обеспечивают высокую компартментализацию организма и выполняют функцию, аналогичную роли этих элементов в составе гистогематических барьеров позвоночных (N. J. Lane, M. J. Chandler, 1980).
Между эпителиоцитами вторичноротых беспозвоночных - иглокожих - располагаются соединительные комплексы, построенные из десмосомальных и септированных соединений. Так, клетки эпидермиса морских ежей, звезд и голотурий связаны септированными соединениями, ширина межмембранного пространства которых равна 20 нм, толщина септ - 8 нм, а промежутки между септами - 15-20 нм. Ближе к апикальной поверхности расположены зоны слипания. Подобные соединительные комплексы обнаружены между эпителиоцитами целома, к которым посредством десмосом прикреплены миоциты. Септированные соединения находятся также между эпидермальными и подлежащими клетками. У морских ежей к септированным соединениям часто прилегают митохондрии (S. Hamada, 1979).
У многих видов иглокожих (морских ежей, звезд и голотурий) обнаружены своеобразные анастомозирующие септированные соединения. Они характеризуются межмембранным пространством шириной 15-18 нм, заполненным анастомозирующими септами, и образуют (вместе с десмосомами) соединительные комплексы. Поскольку строение мембран в этой области напоминает структуру мембран плотного соединения, следует полагать, что анастомозирующие септированные соединения иглокожих выполняют, подобно плотным, барьерную функцию (C. R. Green и др., 1979). Истинные плотные соединения у этих вторичноротых животных до сих пор не обнаружены (P. R. Matzia и др., 1980).
Большой интерес представляет изучение МК в эпителиях щетинкочелюстных - животных, близких по своему строению к представителям вторичноротых. Установлено, что соединительные комплексы между эпителиоцитами кишки сагитты состоят из зон слипания, парных септированных соединений, обычных септированных соединений и щелевых соединений.
Зоны слипания имеют типичное строение и характеризуются межмембранным пространством шириной 13 нм. С помощью этих зон осуществляется объединение апикальных фибриллярных сетей контактирующих клеток.
Парные септированные соединения впервые обнаружены у щетинкочелюстных. В этих соединениях межмембранное пространство шириной 12 нм заполнено септами, которые располагаются парами; каждая пара септ отделена от соседней значительным промежутком. При криоскалывании на Р-поверхности в области парного септированного соединения выявлены гребешки, состоящие из внутримембранных частиц диаметром 10 нм. На Е-поверхности определяются комплементарные бороздки.
Обычные септированные соединения располагаются ближе к базальной поверхности эпителиоцитов и по структуре резко отличаются от парных септированных соединений. Щелевые соединения принадлежат к типу Р, большое количество их у щетинкочелюстных, по-видимому, объясняется отсутствием кровеносной системы у этих животных.
У щетинкочелюстных обнаружены своеобразные соединения, соединяющие три контактирующих эпителиоцита, которые получили название "трехклеточные соединения" (M. Duvert и др., 1980). Они состоят из отдельных субъединиц, а каждая субъединица - из центрального кольца диаметром 15 нм и трех мостиков, соединяющих центральное кольцо с тремя контактирующими эпителиоцитами. Таким образом, МК этой своеобразной группы беспозвоночных животных отличаются значительным своеобразием. Плотные соединения у щетинкочелюстных пока не выявлены. Представляет интерес исследование МК в многослойном кожном эпителии щетинкочелюстных животных.
Иной план строения имеют МК кишечнодышащих. У этих животных наряду с десмосомами обнаружены плотные соединения. В кожном эпителии кишечнодышащих находятся зоны слипания вблизи апикальной поверхности эпителиоцитов (М. В. Атаманова, 1980). В области этих соединений плазмолеммы соседних клеток располагаются параллельно друг другу, цитоплазма содержит значительное количество электронноплотного материала, с которым связаны пучки фибрилл. В месте контакта клетки образуют выступ, вдающийся в цитоплазму соседнего эпителиоцита. Плотные и щелевые соединения, а также десмосомы выявлены в эпидермисе и других эпителиальных тканях оболочников. Важно отметить, что оболочники - единственные представители низших хордовых, у которых наличие плотных соединений доказано с помощью параллельного исследования срезов и сколов (D. Georges, 1980).
Исходя из этих данных можно было бы считать, что у хордовых наиболее характерными должны быть подобные элементы. Однако у ланцетника в МК эпидермиса еще не выявлены десмосомы, плотные соединения и иные специализированные элементы. В частности, отсутствуют септированные соединения. Вместо них межклеточные взаимоотношения обеспечивают неспециализированные простые соединения, протяженность которых значительно увеличивается за счет интердигитаций (E. Schulte, R. Riehl, 1977).
Таким образом, у иолухордовых и низших хордовых структуру МК нельзя представить единой схемой. Можно лишь отметить, что у этих животных септированные и щелевые соединения пока не обнаружены. В противоположность этому МК позвоночных имеют единообразный план строения. Между апикальными областями эпителиоцитов позвоночных располагается типичный соединительный комплекс. Подобные соединительные комплексы, состоящие из трех элементов, наблюдаются в эпителиальных тканях представителей всех классов позвоночных и хорошо выражены уже у круглоротых. С помощью криоскалывания в их составе часто выявляют и щелевые соединения. Так, в щитовидной железе ряда животных щелевые соединения располагаются в составе соединительных комплексов и окружены элементами плотных соединений (K. Ishimura, H. Fujita, 1979, В. P. M. Menco, 1980).
Следовательно, на третьем этапе становления системообразующей функции МК появляются соединительные комплексы, которые характеризуются строго упорядоченным расположением входящих в их состав соединений. У двух филогенетически далеких групп многоклеточных животных - позвоночных и высших беспозвоночных - в соединительные комплексы включаются структурно и генетически различные, но функционально аналогичные соединения (например, плотные и септосодержащие). Появление соединительных комплексов значительно повысило эффективность контактных взаимодействий клеток, увеличило число уровней иерархической организации МК: уровень соединений - уровень комплексов - уровень контактной площадки - уровень контакта (В. И. Архипенко и др., 1975). Однако для образования комплексов потребовалось создание сложной регуляторной системы, координирующей взаиморасположение и взаимодействие отдельных специализированных соединений. Согласно представлениям, развиваемым В. Ф. Ушаковым (1980, 1981), координация взаимного расположения специализированных элементов на контактной площадке может осуществляться с помощью десмосом.
Проведенный анализ свидетельствует о правомерности выделения трех этапов в становлении системообразующей функции МК. Однако многие важные аспекты этого процесса еще нуждаются в исследовании. Недостаточно изучены контактные взаимоотношения клеток у таких важных в эволюционном отношении типов, как губки, плоские и кольчатые черви, а также низшие хордовые. Не исследованы МК форонид, мшанок, плеченогих, погонофор и других типов беспозвоночных животных. Выяснение всех этих вопросов позволит проверить и уточнить предлагаемую периодизацию филогенетического становления системообразующей функции МК.
Третья задача сравнительной контактологии - разработка систематики МК. Одним из наиболее важных является вопрос об аналогии и гомологии отдельных специализированных элементов контакта. Критерии для установления аналогии элементов МК исходят из анализа функций каждого специализированного элемента в соответствии с тремя составляющими системо-образующей функции: адгезией, коммуникацией и изоляцией.
При разработке критериев для установления гомологии специализированных элементов контакта необходимо исходить из результатов анализа структуры каждого элемента в соответствии с теоремой гомологии ультраструктур (R. Rieger, S. Tyler, 1979). В настоящее время известны три критерия гомологии ультраструктур, которые следует применять при изучении сравнительной морфологии МК. В качестве дополнительного, четвертого, критерия гомологии можно использовать способность элементов МК к гибридизации.
В экспериментальных условиях между клетками представителей различных видов, классов и даже типов животных могут образовываться гибридные соединения. Так, между сращенными частями различных гидр образуются щелевые соединения, которые могут передавать морфогенетическую информацию, необходимую для интеграции трансплантата и хозяина (R. J. Wakeford, 1979). В условиях трансплантации клетки различных видов кишечнополостных способны образовывать различные специализированные соединения; описано образование контактных структур между клетками различных видов иглокожих (M. Spiegel, E. Spiegel, 1978).
Гибридные десмосомы формируются в экспериментальных условиях между клетками различных классов позвоночных (J. Overton, 1977; А. С. Nag, M. Cheng, 1980), но пока не удалось добиться формирования полноценных специализированных соединений между клетками позвоночных и животных других типов. Клетки различных видов слизевиков при совместном культивировании рассортировываются и образуют агрегаты из клеток одного вида. Таким образом, образование гибридных МК может быть одним из перспективных методов исследования этих структур. Дальнейшее развитие данного направления позволит получить новую информацию о гомологии элементов контакта у представителей отдаленных систематических групп животных и на этой основе создать эволюционно обоснованную систему специализированных соединений МК.
Особый интерес представляет возникновение в процессе эволюции плотных соединений, которые у низших беспозвоночных не обнаружены. Высказаны две гипотезы о возникновении плотных соединений. Согласно первой, предшественниками плотных соединений явились различные формы септированных, выполняющих, так же как и плотные, барьерную функцию у многих типов беспозвоночных животных (C. R. Green и др., 1979). Однако идентичность изолирующей функции плотных и септированных соединений свидетельствует скорее об их аналогии. Кроме того, не найдены соединения, промежуточные по структуре между плотными и септированными, что является необходимой предпосылкой для вывода о^ их гомологии.
Согласно второй гипотезе, существует гомология между плотными и щелевыми соединениями. Возможно, что для образования плотных соединений у высших беспозвоночных и позвоночных животных были использованы внутримембранные частицы, подобные частицам щелевых соединений (Л. В. Гербильский, 1980). О гомологии этих функционально различных элементов МК может свидетельствовать сходство их структуры, расположения на контактной площадке, а также корреляции в изменениях данных элементов при патологии (Robenek и др., 1980).
Для проверки данных гипотез необходимы дальнейшие, более углубленные исследования, в том числе изучение аминокислотной последовательности белков внутримембранных частиц всех типов специализированных элементов МК у различных животных. При этом необходимо учитывать различия в аминокислотной последовательности белков соединений одного типа в разных тканях (R. J. Nicholson и др., 1980; M. Friedlander, 1980).
Четвертая задача сравнительной контактологии - изучение роли контактных взаимодействий клеток в патологических процессах у животных, находящихся на различных этапах эволюции. В этом направлении получены немногочисленные данные. Однако не вызывает сомнения, что сложно построенные МК позвоночных животных, разделяющие организм на относительно изолированные отсеки, могут подвергаться многочисленным повреждающим воздействиям. Нарушение барьеров между различными компартментами - возможное звено аутоиммунных и других патологических процессов.
Таким образом, повышение системообразующей функции МК, явилось основой для усовершенствования структуры тканей и органов многоклеточных животных и создало предпосылки для развития патологических процессов в этих органах.
Познание закономерностей эволюционного становления системообразующей функции контактов, решение задач, стоящих перед сравнительной контактологией, должно явиться теоретической базой для эволюционно обоснованного управления контактными взаимодействиями клеток измененных органов.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'