Изучение ориентации морских черепах в море и на суше (А. Карр)
В течение последних 6 лет проводилось изучение биологии пяти видов американских морских черепах. Был, наконец, установлен ценный и давно подозреваемый факт миграции черепах на большие расстояния. Однако остается еще ряд других проблем, и их нельзя решить до тех пор, пока не будут разработаны методы непрерывного наблюдения за черепахами при всех изменениях, происходящих в процессе онтогенеза или в экологии черепах, и при их перемещении на большие расстояния. Только применение телеметрической аппаратуры позволит, вероятно, решить ряд неясных вопросов, касающихся их экологического ареала, а также целый комплекс проблем, связанных с ориентацией - от простых таксисов до ориентации по небесным светилам и без видимых ориентиров.
В одной из работ, у Тортугуеро (Коста-Рика) на берегу Карибского моря, было проведено мечение 2000 взрослых самок зеленой черепахи Chelonia. После сезона кладки яиц наблюдалось 64 случая возвращения меченых черепах, причем 62 особи были пришельцами из других мест. Эти черепахи были замечены в различных участках всего западного берега Карибского моря. Полученные данные свидетельствуют в пользу того, что периодическая миграция черепах действительно происходит, но строгих доказательств получено не было. Основная часть популяций зеленых черепах возвращается для откладки яиц каждые 3 года, но, как показали их повторные появления на берегу Коста-Рики, 30-40% из них имеет дополнительный двухгодовой цикл. Мы много раз ловили у Коста-Рики черепах, которым была нанесена метка 2-3 года назад. Так как на побережье Коста-Рики черепахи не живут, эти возвращения подтверждают сведения о том, что черепахи мигрируют между Тортугуеро и отдаленными районами, где они кормятся. Приверженность к определенным местам гнездования характерна не только для черепах, возвращающихся после длительного отсутствия, но и для черепах, возвращающихся 3-4 раза во время одного сезона для откладки яиц.
Чтобы приблизиться еще хоть на дюйм к прямому доказательству миграции черепах, мы предприняли мечение черепах на острове Вознесения в южной части Атлантического океана. В этом месте черепахи откладывают яйца только в определенное время года, полностью исчезая, когда откладка яиц окончена. Черепахи, постоянно обитающие на побережье Бразилии, никогда не откладывают яиц на этом побережье. Нам казалась поэтому правдоподобной гипотеза о том, что бразильские черепахи откладывают яйца на острове Вознесения. Весной 1960 г. было проведено мечение 206 самок на острове Вознесения. То, что все 5 вернувшихся до настоящего времени были с бразильского берега, доказывает нашу гипотезу, насколько это возможно при простом мечении. Но даже эти скромные результаты обесцениваются тем, что мы совершенно не знаем маршрутов и графиков плавания черепах и способов их ориентации. Добиться лучшего удастся лишь в том случае, если будет возможно непрерывное прослеживание мигрирующих черепах.
Использование баллонов с гелием для прослеживания
Было установлено, что после длительных морских плаваний черепахи возвращаются на вполне определенный участок побережья. Поэтому зеленая черепаха представляет собой очень ценный объект для изучения способов ориентации животных. Первые предварительные опыты прослеживания были выполнены на трех видах черепах; к панцирям прикрепляли поплавки из пенопласта, к которым были привязаны баллоны, наполненные гелием (фиг. 1). Восемь черепах, закончивших откладку яиц, были выпущены на отмель с целью изучения их поведения в 12-дневный период до следующей откладки яиц. Остальные опыты проводились в открытом море вдали от берега. Прослеживание проводилось при помощи теодолитов, алидад и дальномеров. Большие трудности в методах измерения приводили к двусмысленным и ошибочным данным, что обесценивало значительную часть работы. При опытах в прибрежных водах прибой затруднял даже доставку оборудования. Потребовалось несколько испытаний для определения глубины погружения черепахи и длины троса, при которой поплавок остается на поверхности. Выяснилось, что невозможно предусмотреть неожиданное погружение черепахи, когда она замечает впадину в скале или другое место, где находится корм.

Фиг. 1. Приспособление, используемое в опытах прослеживания с баллонами. Поплавок из пенопласта, который держит человек слева, прикреплялся к задней части панциря черепахи при помощи 9-метрового многожильного троса. К поплавку прикреплялись два баллона на тросах длиной 3 и 4,5 м. Шарнирное соединение, к которому был прикреплен трос, присоединялось к панцирю тонкой железной проволокой. Через некоторое время эта проволока ржавела и ломалась, освобождая черепаху от приспособления. Всех подопытных черепах метили стандартными метками на конечностях
Были и другие трудности. Так, при длительных опытах улетучивался гелий или баллоны перегревались и взрывались. Многие баллоны лопались, когда поднимался легкий бриз или трос цеплялся за пловучие водоросли. Использовать для баллонов водород было невозможно, так как его отказывались перевозить капитаны судов.
Недостаток визуального прослеживания состоит в том, что его нельзя проводить ночью. Можно было бы, конечно, прикрепить к баллону источник света, но такое "украшение" ночного неба, видимого черепахой, нарушит постановку опыта. Вполне вероятно, что черепахи замечают баллоны и днем, и это как-то влияет на опыт, однако освещение баллона ночью оказало бы значительно большее влияние.
Все это указывает на то, что для качественного прослеживания необходима телеметрия. Как уже говорилось, весьма подходящий объект для таких исследований представляют собой морские черепахи Chelonia, особенно взрослые самки, вес которых составляет обычно 112,5-157,5 кг. К выступающим краям панциря этих больших и сильных животных легко прикрепить болтами аппаратуру, не вызывая излишнего раздражения ткани, причем миниатюризации приборов не требуется. Удобно также то, что скорость черепахи сравнительно мала - не более 32-48 км в сутки при непрерывной миграции. Вода плохо проводит радиоволны, но черепаха дышит воздухом и должна подниматься на поверхность каждые 1-3 мин при движении и 2-30 мин при покое. При вдохе передний край панциря поднимается до уровня воды или даже на несколько сантиметров выше. В этом отношении черепаха - менее удобный объект для радиопрослеживания, чем птицы, но гораздо более удобный, чем рыбы.
Изучение ориентации черепахи Chelonia имеет косвенное отношение к проблеме определения направления по солнцу. В открытом море нет неподвижных ориентиров, необходимых для нахождения азимута. Поэтому предположение об ориентации по солнцу кажется маловероятным. По мнению Пенникуика [4], птицы могли бы определять свое положение либо по высоте солнца, либо по скорости изменения высоты. Опыты Шмидта-Кёнига [5] дают основания сомневаться в том, что птицы действительно используют этот способ. Если принять, что черепахи ориентируются по солнцу, то придется предположить, что они используют в качестве вспомогательного ориентира направление движения волн. Однако черепахи ориентируются как над водой, так и под водой. Следовательно, направление движения волн не играет никакой роли при их движении. В настоящее время у нас еще нет теории, объясняющей искусную ориентацию черепах при дальних морских плаваниях.
Трудности прослеживания черепах
Кроме поддержания непрерывного контакта с черепахой, важно знать, что она пытается делать, куда она, так сказать, стремится. Если цели некоторых движений черепах можно предугадать, то другие движения (не менее интенсивные) осуществляются безо всякой заметной цели. Хорошим примером служит беспорядочное движение молоди. Только что вылупившиеся черепахи, помещенные в резервуар, непрерывно и энергично плавают вдоль и поперек него. Если резервуар с одной стороны освещен, то черепахи собираются к свету, но и при этом они продолжают свое бесцельное плавание в течение дней или даже недель. Это плавание, возможно, ничего не означает, но оно помогает объяснить полное исчезновение молодых морских черепах всех видов по крайней мере на год после того, как они впервые пересекают линию прибоя. Результаты этих наблюдений позволяют предположить, что молодые черепахи непрерывно плавают в течение длительного периода, чем и объясняется их исчезновение. Прослеживание животных в этот период позволило бы заполнить существенный пробел в исследовании полного жизненного цикла черепах. Описанные выше опыты с освещенным резервуаром указывают, возможно, также на то, что латентная способность к ориентации проявляется у молодых черепах после завершения ряда событий, связанных с их передвижением от гнезда к линии прибоя. Однако эксперименты мало что смогут добавить к этому, пока прослеживание не покажет, что "чувство компаса" заложено в природе черепахи и не поможет предсказать направление ее движения.
Непредсказуемость движений черепах является наибольшим препятствием при прослеживании их на небольших расстояниях. Об этом свидетельствуют результаты шести опытов прослеживания, проведенных у Кедровой отмели (западное побережье Флориды). Для прослеживания были выбраны взрослые самки черепах вида ложная каретта (Caretta caretta), направлявшиеся к месту откладки яиц. Такой выбор был обусловлен тем, что при этом виде миграции морские черепахи движутся к своей цели исключительно настойчиво. Мы наблюдали, как около нашего лагеря на побережье Коста-Рики черепахи выходили на сушу, волоча за собой прикрепленные к ним большие буи, по которым мы их отыскивали в море. Кроме того, как указывалось выше, черепахи проявляют большое постоянство в выборе места для откладки яиц. При нескольких откладках яиц в один сезон они возвращались на один и тот же небольшой участок 34-километрового берега или в одну и ту же бухту на одном острове [2] или на один и тот же островок из группы в несколько небольших островков, расположенных поблизости друг от друга [3].
Шесть опытов у Кедровой отмели носили разведочный характер, и вскоре мы убедились, что они были поставлены неправильно. Черепахи выпускались так, что полуостров Флорида находился между ними и их предполагаемой целью. Это весьма усложняло их навигационные задачи. Более логичным было бы, конечно, выпускать черепах в Атлантическом океане. Район Кедровой отмели был выбран по той причине, что линия дневных маяков вдоль рифов помогала при оптических наблюдениях. В океане эти наблюдения пришлось бы проводить с небольших лодок и, вероятно, при сильном волнении.
Правильная или "бессмысленная" навигация
При проведении подобных исследований нужно дать ответ на следующие три вопроса: 1) движется ли животное целенаправленно или случайно, 2) движется ли оно по направлению к дому, 3) попадает ли оно домой и если да, то по какому пути (т. е. приводит ли этот путь, если его экстраполировать и внести некоторые поправки, к дому. В противном случае навигация будет, по выражению Мэтьюза, "бессмысленной").
В наших опытах на Кедровой отмели невозможно было сказать заранее, какое из направлений движения является правильным. Как могла, например, ориентироваться морская черепаха с берега Дейтоуна, которая была поймана, перевернута на спину и перевезена ночью в крытом грузовике на противоположный берег полуострова Флорида? Она могла просто счесть, что находится "в открытом море", и направиться к определенному каким-то образом ближайшему берегу. Или, исходя из того же предположения и действуя более сложным образом, она могла направиться на запад в том направлении, в каком ей нужно было направляться к берегу из Атлантического океана. Если допустить, что черепаха хорошо ориентируется по звездам, но не имеет врожденного знания географии земли, то она могла бы направиться на родной берег, т. е. через полуостров. Возможно также, что черепаха, так же как и некоторые птицы, обладает самыми совершенными способностями к ориентации (врожденное чувство звездного календаря и земной географии) и либо направится к югу, чтобы обогнуть полуостров, не видя его берегов, либо подойдет к берегу, а затем повернет к югу, следуя вдоль берега. Очевидно, предсказать заранее, какой путь она выберет, невозможно. Нельзя также проводить уверенную экстраполяцию по небольшим отрезкам пути черепахи. Сочетание этих неопределенностей усложняет опыты с морскими черепахами, проводимые в прибрежных водах.
Суммарные результаты шести опытов не показывают совпадения первоначальных направлений движения. Однако можно предположить, что некоторые черепахи выбирали направление не случайно. В частности, графики движения 3 черепах, приведенные на фиг. 2 и 3, показывают, что две черепахи выбрали направление на юг. Их местонахождение определялось каждые две минуты, что позволяет считать графики абсолютно точными.
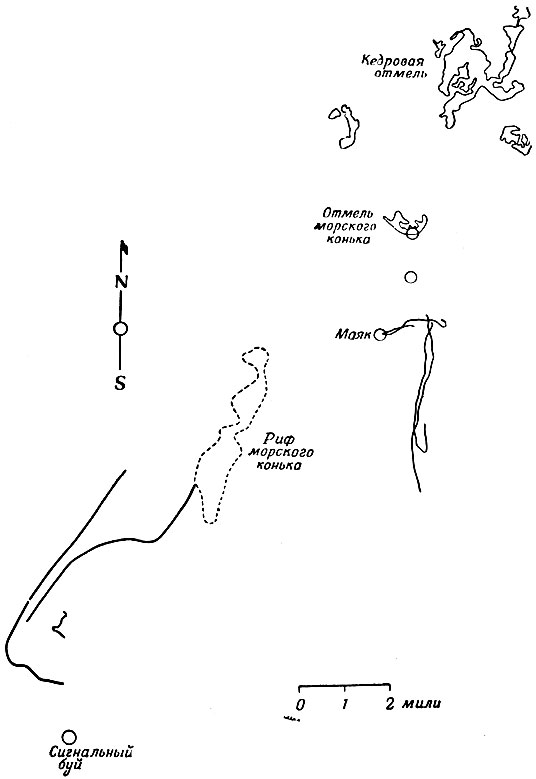
Фиг. 2. Результаты прослеживания самок черепахи вида Caretta caretta на Кедровой отмели у побережья Флориды (1960). Черепах перевезли через полуостров от форта Пирс, где они собирались отложить яйца. Пути трех черепах в левой части фигуры получены при шестичасовом прослеживании с помощью двух алидад, одна из которых находилась на маяке, а другая на лодке. Отрезки пути трех других черепах в правой части фигуры (см. также фиг. 3) получены при помощи двух теодолитов, расположенных на расстоянии 1,85 км на дневных маяках. Два южных курса получено при прослеживании 9 июля, третий - 26 июня
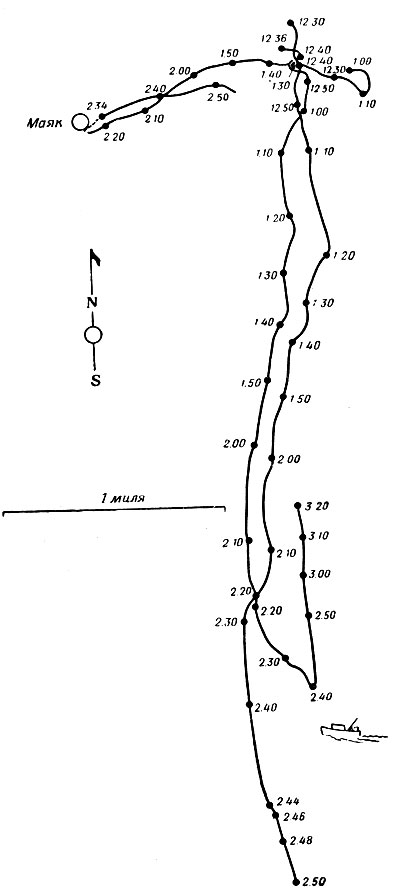
Фиг. 3. Результаты прослеживания трех черепах вида Caretta caretta у Кедровой отмели (см. фиг. 2). К черепахам были прикреплены гелиевые баллоны. Прослеживание проводилось при помощи двух теодолитов. Положение черепах отмечалось каждые 2-3 мин
Так как графики ничего не говорят о причинах выбора направления, вопрос о том, случайно ли движение черепах, по-прежнему остается открытым. Все время пути они находились вдали от материка, так что он был для них невидим, и из-за малого угла наблюдения почти не видели никаких неподвижных ориентиров. Судя по карте, черепахи вряд ли могли бы ориентироваться по рельефу морского дна. Курс пролегал через воды различного химического состава, и морские течения менялись по силе и направлению. На значительных отрезках пути черепахи не видели друг друга (каждая из них могла видеть лишь баллон, привязанный к другой черепахе, но трудно предположить, что это означало какую-либо связь между ними), и это позволяет еще более высоко оценить совершенство их навигации. Единственным возможным ориентиром был довольно неподвижный ряд кучевых облаков над материком (на расстоянии 24-32 км). Маловероятно, чтобы черепахи не использовали этот ориентир, но вряд ли они точно выдерживали направление движения на юг только благодаря ему. То же самое можно сказать об ориентации по солнцу на тех небольших расстояниях, которые проплыли черепахи за время опыта.
Планы будущей работы
Опыты с баллонами позволили наметить программу дальнейших исследований. Прежде всего, необходимо узнать, могут ли черепахи находить определенное направление, не используя видимые ориентиры. О том, что они способны к этому, свидетельствует их географическое распространение. Но опыты с баллонами, по-видимому, не смогут дать достаточно надежной подтверждающей информации. Очевидно, что это как раз тот случай, когда телеметрия может оказаться весьма полезной.
Проведенные исследования показали способность черепах ориентироваться в открытом море. Дальнейшие опыты по выяснению механизмов ориентации можно разделить на две части: 1) изучение ориентации черепах в знакомой местности и 2) изучение их ориентации при определении своего местоположения или при возвращении к родному берегу. Именно в этой, второй части исследований особенно необходимо применение телеметрии. Пути, по которым перемещаются черепахи, нам неизвестны. Черепахи, вылупившиеся из яиц на острове Вознесения, и взрослые черепахи, возвращающиеся с этого острова на бразильский берег, могли бы пассивно дрейфовать, используя морские течения, пока они не окажутся вблизи материка. Однако взрослые черепахи, направляющиеся от материка к маленькому острову в океане, могли бы выбрать по меньшей мере три направления движения: 1) движение по течению Гольфстрим, 2) движение по схеме: Бразильское течение - течение Западных ветров - Бенгельское течение, 3) движение напрямик поперек трех ветвей Южного Экваториального течения. Последнее кажется наиболее вероятным, но это движение представляет собой в высшей степени трудную навигационную проблему, если учесть смещения, вызываемые течениями.
На карибском побережье мы встречали черепах только трех возрастов. Взрослые самки приплывали на побережье Коста-Рики на откладку яиц. Их сопровождали взрослые самцы, оплодотворяющие яйца перед откладкой. Там же встречались черепахи, только что вылупившиеся из яиц. В центральной части морского побережья Флориды мы наблюдали молодых неполовозрелых (4,5-40,5 кг) зеленых черепах, которые появлялись в апреле и исчезали в конце октября. Вся остальная часть жизненного цикла остается неизвестной. Следовательно, предсказание направления движения возможно лишь для черепах немногих определенных возрастов, и оценивать направление движения по коротким отрезкам пути следует с величайшей осторожностью.
Только в одном-единственном случае нетрудно изучить ориентацию черепах, хотя механизм ее и нелегко понять. Этим случаем является короткое, но полное событий путешествие только что вылупившихся черепах от гнезда к морю. Очевидно, что при этом они не определяют свое местонахождение относительно стран света. Черепахи, вылупившиеся из яиц, взятых на побережье Атлантического океана (океан находился к северу от берега) и перенесенных на побережье Тихого океана (океан находился на юго-западе от берега), вели себя так же, как на родном берегу, и столь же успешно достигали воды. Тем не менее отважные поиски моря только что вылупившимися черепахами представляют собой хорошую модель сложной и искусной ориентации взрослых черепах. Основной реакцией при поисках моря является один из видов телотаксиса, а именно стремление двигаться в направлении более освещенной части неба или к горизонту, свободному от облаков. Но по пути этот ориентир дополняется или заменяется другими, т. е. происходит непрерывная переориентация. Изучение путешествия молодых черепах позволяет проникнуть в природу инстинктивной ориентации и укрепляет уверенность исследователей в том, что ориентация в пути всегда слагается из нескольких отдельных процессов.
Путь только что вылупившихся черепах к морю
Путь к морю начинается под слоем песка толщиной примерно 70 см (фиг. 4); вылупившиеся из яиц черепахи оказываются как-бы в камере, так как в результате опадания яйцевых оболочек в песке образуется некоторое свободное пространство. Доказано, что путь на поверхность - это не просто движение отдельных особей сквозь песок под влиянием отрицательного геотаксиса, как предполагалось ранее. Стремление пробиться вверх играет, конечно, некоторую роль, но основным фактором, определяющим выход на поверхность, является движение вверх всего скопления молоди, представляющее собой некоторый первичный вид сотрудничества. "Пусковым механизмом" этого процесса служит активность особей, находящихся на дне камеры. Под влиянием этой активности начинается бурная деятельность всей массы вылупившихся черепах. Они подкапывают стены, скребут потолок или утаптывают песок на дне камеры, которая постепенно поднимается вверх. Групповая деятельность автоматически переходит в некоторое подобие разделения труда, в результате которого вся камера с выводком перемещается вверх. Черепахи, вылупившиеся на той же глубине поодиночке, очень часто погибают.

Фиг. 4. Подъем выводка черепах вида Caretta caretta. Первоначальное расположение выводка можно определить по спавшимся яйцовым оболочкам у дна ящика. Там же осталось несколько аномальных особей. Остальные 65 молодых черепах объединенными усилиями пробиваются на поверхность (см. текст)
Движение выводка на поверхность продолжается около недели. При наблюдении этого процесса через стеклянную стенку естественная картина несколько искажается, так как на животных действуют такие посторонние факторы, как сильный свет, требуемый для наблюдения, и наличие твердой стенки. Для изучения этого интереснейшего случая взаимопомощи необходимы приборы, которые позволили бы наблюдать положение и деятельность каждой из черепах в течение всего периода выхода на поверхность.
После появления выводка на поверхности основная его задача состоит в отыскании воды, которая чаще всего скрыта за дюнами, кустами, пловучим лесом, прибитым к берегу, и т. п. (фиг. 5-7). Черепахи осматриваются, начинают двигаться и после небольшого периода ошибок почти всегда поворачивают прямо к океану. Они карабкаются по склонам, обходят поваленные деревья, проползают под кустами и через мусор, выброшенный на берег. Кажется, что все они, кроме аномальных и слабых особей, твердо знают некий маяк, указывающий им путь к океану. Они находят океан днем и ночью. Дождь, облачность, освещенность местности или океана не играют при этом никакой роли.

Фиг. 5. Сложный профиль морского берега, в условиях которого выводки черепах успешно ориентируются и находят путь к морю (гнездо показано кружком слева)

Фиг. 6. Две молодые зеленые черепахи, успешно ориентирующиеся на местности после выпуска их на краю дюны у берега Дайтона (Флорида). Эти черепахи, вылупившиеся из яиц у Тортугуеро (Коста-Рика), взяты из выводка, находившегося 9 месяцев в 7-литровых банках в лаборатории. Они никогда не видели моря. Но они шли прямо к морю, преодолевая промоины и крутые песчаные валы, заметные на заднем плане фотографии

Фиг. 7. Группа молодых черепах, идущих в направлении моря (на фотографии - вверх направо). Близкое белое пятно привлекало внимание черепах, но лишь на несколько секунд
Несмотря на большое количество работ различных исследователей (см. обзор в работе Карра и Огрена [1]), посвященных морским и пресноводным черепахам, основной ориентир черепах в пути остается неясным. Какое-то значение имеет освещенность неба над морем. Если бы это была простая положительная реакция на свет - простой фототаксис, - то можно было бы ожидать, что выводок будет направляться в сторону солнца или луны; однако это случается очень редко. Вместе с тем, если черепаха сначала находится за препятствием, а затем в ее поле зрения попадает более яркое небо над морем, то она сразу же начинает двигаться быстрее. Черепахи сразу устремляются вперед, когда замечают ночью белую под лунным светом полосу прибоя или яркий бурун в море. Но даже при полном штиле черепахи редко ошибаются в отыскании пути к океану, где бы ни находилось их гнездо. Иногда случаются и большие ошибки, но они редки и сами по себе заслуживают изучения.
После трудного пути по сыпучему песку дюн черепахи достигают плотного слоя песка у линии прибоя. Здесь, кроме яркой линии прибоя, появляются и другие ориентиры - по-видимому, твердая, ровная и наклонная поверхность. Черепахи сразу начинают двигаться быстрее; паузы для осмотра местности становятся все более редкими.
Затем достигается влажный песок, куда докатываются самые высокие волны. Здесь наблюдается новый скачок скорости и уверенности движения. Некоторые черепахи как бы вдруг спотыкаются и начинают преждевременные плавательные движения. Прикосновение влажного песка служит, вероятно, сигналом к появлению иной "походки", которая окажется полезной только при достижении воды.
Самая дальняя волна накатывается на песок и поднимает на пол сантиметра находящихся впереди черепах. Как только они отрываются от земли, они начинают энергичные плавательные движения. Когда волна уходит, они продолжают делать эти движения, мало пригодные для ходьбы на суше. Каждая новая волна немного продвигает их вперед к линии прибоя.
Такие плавательные движения представляют собой, по-видимому, безусловный рефлекс и обучаться им специально черепахи не должны. После того как волна подымет их несколько раз, они уже "обучены", причем сразу же проявляется биполярный реотаксис, помогающий им подстраиваться к уходу и приходу волн. Очевидно, случайное прикосновение ко дну подсказывает черепахам, что вода находится в движении и что она перемещается то в одну, то в другую сторону.
Так или иначе, черепахи держат курс прямо в море. Они энергично проплывают под водой 1-1,5 м, затем поднимаются на поверхность, чтобы сделать вдох и осмотреться, снова уходят под воду и появляются на поверхности. Когда черепахи достигают буруна, то кажется, что он поднимет их и отбросит назад через весь пройденный путь. Но вот вырастает белый гребень, и головы черепах исчезают. Вероятно, выводок ныряет на дно и плывет под гребнем. Конечно, этот момент путешествия к морю полностью скрыт от наблюдателя, но можно с уверенностью сказать, что при перемещении черепах у дна они проявляют целый ряд реакций на движение волн. Поведение выводка под волной прибоя нам неизвестно, но, без сомнения, черепахи прекрасно приспосабливаются к динамике волн. Если бы мы могли осуществить непрерывное наблюдение за этим процессом, то это не только пополнило бы наши знания об одном из интереснейших механизмов ориентации, но и пролило бы свет на необъяснимое отсутствие морских черепах на тысячекилометровых побережьях тропических морей.
Таким образом, совершая короткое, но сложное путешествие от гнезда к морю, черепахи направляются к цели, которую они никогда не видели, и используют при этом информацию от целого ряда источников. Так же как и в случае изучения миграции на большие расстояния, одна лишь оценка ориентиров не в состоянии объяснить, как устанавливается и соблюдается любой длительный курс животных и куда он ведет. Идеальным методом таких исследований явилось бы радиопрослеживание, которое позволило бы непрерывно следить за положением мигрирующих черепах в течение всего их пути и возращения на родной берег. То, что о таком идеале можно говорить всерьез, свидетельствует о наступлении новой эпохи в развитии естествознания.
Обсуждение доклада
Бах. Обсуждая этот доклад, следует вспомнить о работах д-ра Баллета. Я не знаю, опубликовал он их или нет. Опыты проводились в Калифорнии, где пустыня подходит к морю. Баллет взял вылупившихся из яиц морских черепах, которые никогда не видели пустыни, и выпустил их на склоне горы, примыкающем к пустыне. Живущих в пустыне пресмыкающихся, которые никогда не видели моря, он перевел на склон горы, обращенной к морю. Животные направились в противоположные стороны извернулись к родным местам. Я не знаю, как, но они сделали это.
Корсон. Все ли черепахи в конечном счете приходят к морю? Не теряются ли некоторые из них?
Карр. Небольшая часть черепах пропадает. Обычно это те, которые застревают на берегу и быстро обезвоживаются под действием солнечных лучей. Иногда мы не могли отличить потерю ориентации от физиологической неспособности продолжать движение.
Конечно, некоторые вылупившиеся из яиц черепахи ведут себя аномальным образом. Они не присоединяются к группе, пробивающейся на поверхность, и иногда так и остаются в песке. Некоторые из них не доходят до океана. Несколько раз мы наблюдали необъяснимую ошибку в ориентации черепахи на 180°.
Шмидт-Кёниг. Появляются ли черепахи на поверхности в какое-то определенное время дня?
Карр. Обычно это происходит рано утром, как раз перед рассветом, но не всегда. Вообще возможен выход выводка на поверхность в любое время суток. Это подтверждается также тем, что черепахи ориентируются по отношению к морю в любое время дня или ночи. Их способности к ориентации не зависят от погоды. День или ночь, дождь или ясное небо, солнце или луна над океаном или над сушей - все равно они в большинстве идут прямо к океану.
Шмидт-Кёниг. Может быть, их привлекает шум прибоя?
Карр. Я не думаю. Правда, мы располагаем лишь негативными доказательствами. Нам кажется, что основным фактором является свет. С одним закрытым глазом черепахи двигаются с небольшими поворотами по кругу. Если закрыть им оба глаза, то они вообще теряют свойство определять направление к морю.
Вопрос с места. Вы не пытались проделывать опыты с зеркалом? Я имею ввиду освещение молодых черепах солнечным зайчиком.
Карр. Мы делали это, и черепахи теряли ориентацию. Два раза мы брали с собой на берег большое зеркало и освещали им выводок, идущий к воде. Черепахи немедленно теряли ориентацию.
Вопрос с места. Они не шли к зеркалу?
Карр. Нет. Я предполагаю, что мы просто ослепляли их. Но они могут иногда идти ночью к газолиновой лампе.
Маллинс. Вопрос касается выводка, отвлеченного на короткое время белой маской (см. фиг. 7). Возможно, животные прошли мимо маски, так как она была неподвижна? Я хотел бы знать, могут ли черепахи как-нибудь реагировать на свет, отраженный морем. Я имею в виду то, что волны как-то модулируют отраженный солнечный свет. Этим можно объяснить также то, что черепахи шли на свет лампы.
Карр. Пожалуй, это возможно. Я не знаю, что отличает свет, отраженный океаном, от света фонаря. Дело, вероятно, не в поляризации. Я просто не знаю, имеется ли какое-нибудь изменение в свете типа мерцания, которое они могли бы видеть. Но это возможно.
Мак-Леннан. Будем ли мы рассматривать возможности применения сонарной техники для прослеживания черепах?
Карр. Опыты с баллонами позволяют вести прослеживание на расстоянии до 24 км. Если подвергнуть результаты статистической обработке, то это даст возможность определить, ориентируются ли черепахи. Обеспечит ли сонар большую дальность прослеживания?
Очень ценные сведения дало бы прослеживание черепах, мигрирующих к острову Вознесения. Проплыв почти 2000 км, черепахи уверенно находят десятикилометровую цель в открытом океане. Они явно ориентируются, причем здесь нет никаких неподвижных ориентиров и даже речи о них быть не может. Но как проследить это путешествие?
Выступление с места. Вы бы удивились, узнав, как совершенны некоторые сонары. На противоположных сторонах панциря можно прикрепить передатчики с воздушным отражателем между ними и таким образом определить, какое направление выбрала черепаха.
Галлер. Вы, вероятно, знаете, что были разработаны радиомаяки для прослеживания морских животных, причем был построен опытный экземпляр. Эти маяки могли бы быть полезны в некоторых из ваших опытов с черепахами.
Хаан. Я хотел бы спросить д-ра Карра, слышат ли черепахи те же звуки, что слышим мы, и воспринимают ли они более низкие или более высокие частоты. Не может ли использование сонара внести искажение в те результаты, которые мы хотим получить?
Карр. Вероятно, может. К сожалению, я ничего не знаю о слухе морских черепах. Не следует игнорировать ту возможность, что две черепахи, у которых был одинаковый южный курс (см. фиг. 2 и 3), поддерживали между собой непрерывную связь звуком. Зрительной связи между ними не было - они не видели друг друга. Но я не верю, что их навигационные способности достаточно хороши для точного воспроизведения одинаковых изгибов курса на большом расстоянии. Мы не должны исключать возможность того, что они издают какой-то писк или ворчанье и используют его при совместном плавании.
Хаан. На меня произвел большое впечатление тот факт, что самолет может сбросить в море сонарный блок и получать от него сигналы на расстоянии 2000 км.
Галлер. Я хотел внести кое-какие разъяснения в вопрос о сонаре. Военно-морской флот действительно имеет портативное сонарное оборудование, которое может обеспечить прослеживание на расстоянии до 2000 км, но я подозреваю, что для него нужна платформа размером по крайней мере с миноносец.
Кочрен. Я уверен, что если на панцирь черепахи прикрепить передатчик, описанный д-ром Зингером или д-ром Маршаллом, то мы получим радиус действия 32- 48 км при условии, что черепаха достаточно часто поднимается на поверхность. Как известно, радиоволны хорошо распространяются над поверхностью моря. Если к черепахе можно прикрепить баллон, то она могла бы нести и антенну.
Вопрос с места. Как высока должна быть антенна?
Карр. Черепахи плывут довольно глубоко. Но они дышат, и поэтому не следует беспокоиться о том, что они не выходят на поверхность. Иногда антенна может показаться на гребне волны, а в другой раз - между волнами.
Толлес. Образцовых записей сигналов получить не удастся, но можно будет определить положение черепахи всякий раз, когда она всплывает.
Гриффин. Задача была бы намного проще, если бы антенну можно было прикрепить к баллону.
Литература
1. Carr A., Ogren L., The ecology and migrations of sea turtles, 4. The green turtle in the Caribbean Sea, Bull. Amer. Mus. Nat. Hist., 121, 48 pp., 26 figs., 7 pis. (1960).
2. Carr A., Hirth H., The ecology and migrations of sea turtles, 5. Comparative features of isolated green turtle colonies, Amer. Nus. Novitates, 8/2091, 42 pp., 21 fig. (1960).
3. Hendrickson J. R., The green sea turtle, Chelonia mydas (Linn.) in Malaya and Sarawak, Proc. Zool. Soc. London, 130, pp. 455-535, 15 figs., 10 pis. (1958).
4. Pennycuick C. J., The physical basis of astro-navigation in birds, Journ. Exp. Biol., 37, pp. 572-593, 10 figs. (1960).
5. Schmidt-Koenig K., Sun navigation in birds? Nature, 190, pp. 1025-1026 (1961).
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'