Раздел I. Морфология и физиология риккетсий
Глава 1. Морфология и морфогенез риккетсий
В литературе прочно утвердилось представление о риккетсиях как группе плеоморфных, грамотрицательных, бактериоподобных микроорганизмов, паразитирующих у различных членистоногих, млекопитающих и птиц. Морфологическая характеристика риккетсий, данная на основании изучения фиксированных препаратов из инфицированных тканей, укладывается в схему, предложенную в 1930 г. da Rocha - Lima, определившего, что риккетсии имеют размеры порядка 0,2-0,3×0,3-1 мк, обладают плеоморфизмом, обнаруживаются в виде кокковидных, бациллярных (гантелевидных), палочковидных и нитевидных форм. Однако da Rocha - Lima не связывал существование различных форм риккетсий с цикличностью их развития. Wolbach, Todd, Palfrey (1922) в результате детального изучения морфогенеза риккетсий в кишечнике вшей, зараженных на сыпнотифозных больных, различали три основные формы риккетсий. Первая форма - кокковидные, одиночные и биполярные риккетсии, наиболее часто встречающиеся у вшей; окрашиваются по Романовскому в розовый цвет. Одиночные риккетсии имеют размеры 0,3-0,4 мк, а биполярные - 0,3×0,9 мк. Соединительная часть биполярных форм (гантелей) окрашивается в голубой цвет. Вторая форма - более крупные палочковидные риккетсии длиной 2,5 мк, с четко окрашенными по Романовскому в красный цвет полярными зернами размером 0,25×0,35 мк. Вироплазма риккетсий окрашена в голубой цвет. Третья форма - нитевидные риккетсии, имеющие вид изогнутых или переплетающихся нитей размером 0,3-0,4×10-40 мк. Окрашиваются по Романовскому, как и палочковидные риккетсии. У нитевидных риккетсий нередко отчетливо видна фрагментация, позволяющая различать отдельные палочковидные риккетсии, соединенные в длинную цепочку и объединенные общей оболочкой. Анализируя взаимосвязь описанных форм риккетсий, авторы полагают, что нитевидные формы, возможно, соответствуют наиболее ранней фазе развития возбудителя, тогда как мелкие коккобациллы представляют конечную стадию деления риккетсий.
Весьма обстоятельные исследования по морфологии риккетсий и их размножению выполнены Sikora (1940), которая дала морфологическую характеристику риккетсий, совпадающую с изложенной выше, и выделила два цикла развития риккетсий: а) сокращенный цикл, когда кокковидные риккетсии через промежуточные биполярные формы превращаются в гантелевидные образования, распадающиеся на две самостоятельные особи; б) полный цикл, при котором кокковидные риккетсии превращаются в биполярные, в свою очередь делящиеся с образованцем четырех зернышек. В конечном итоге из одной особи образуются четыре дочерние риккетсии. Выявленный характер размножения риккетсий позволил Sikora отнести их к бактериям.
Сравнительное изучение морфологии риккетсий при экспериментальном риккетсиозе морских свинок, белых мышей, кроликов и развивающихся куриных эмбрионов, проведенное в лаборатории, руководимой П. Ф. Здродовским, позволило дать обобщающую характеристику морфологии и морфогенеза риккетсий. Так, П. Ф. Здродовский отметил чрезвычайно резко выраженный плеоморфизм риккетсий, зависящий от фазы инфекции и ее интенсивности, и выделил четыре морфологических типа риккетсий: a, b, c и d. Тип a, или кокковидные риккетсии, имеет овоидную форму (коккобациллы) и диаметр меньше или около 0,5 мк, часто образует диплоформы, изредка цепочки, а иногда колонии музеровского типа. Эта форма типична для всех видов интенсивного риккетсиоза. Тип b, или палочковидные риккетсии, размером около 1 - 1,5 мк, характерен для интенсивного размножения риккетсий в тканях экспериментальных животных или развивающихся куриных эмбрионов. Тип c, или бациллярные риккетсии, имеет удлиненную и обычно изогнутую форму, размер до 3 - 4 мк. Данный тип характерен для начального периода развития возбудителя и инфицирования малоактивной культурой. Тип d или нитевидные риккетсии, встречается в виде длинных, часто причудливо изогнутых нитей размером 10 - 20 - 40 мк и больше. Эта форма риккетсий, как правило, встречается в случае раннего и умеренного риккетсиоза.
Таким образом, в отличие от других исследователей П. Ф. Здродовский выделил четыре морфологических типа риккетсий, увязывая определенный тип формы и строения со стадиями развития возбудителя. Автор подчеркивает, что более мелкие риккетсии (типа a и b) соответствуют наиболее интенсивному риккетсиозу и наблюдаются в период максимального размножения возбудителя. Бациллярные (c) и нитевидные (d) формы риккетсий характерны для ранних стадий развития в клетках, а также для всех стадий воспроизведения возбудителя при скудном росте риккетсий.
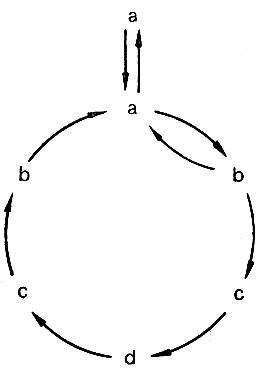
Рис. 1. Схема цикла развития риккетсий. a, b, c, d - формы риккетсий
Что же касается зависимости формы и размеров риккетсий от стадий и способа воспроизведения, то П. Ф. Здродовский доказал существование такой зависимости, установив тесную связь между описанными формами. Схематически динамика перехода одних форм риккетсий в другие в процессе развития может быть представлена в виде замкнутого цикла (рис. 1). Наиболее отчетливо этот цикл прослеживается при репродукции риккетсий Провачека в желточных оболочках куриных эмбрионов, когда удается проследить фазы развития возбудителя на разных этапах его воспроизведения (рис. 2). Действительно, длинные палочковидные и нитевидные риккетсии, имеющие 2 - 4 и более полярных гранул, можно представить как цепочку кокковидных особей, объединенных единой оболочкой. В процессе развития отчетливо прослеживаются сегментация и деление нитевидных форм с образованием кокковидных. В свою очередь кокковидные формы дают начало бациллярным и палочковидным формам.
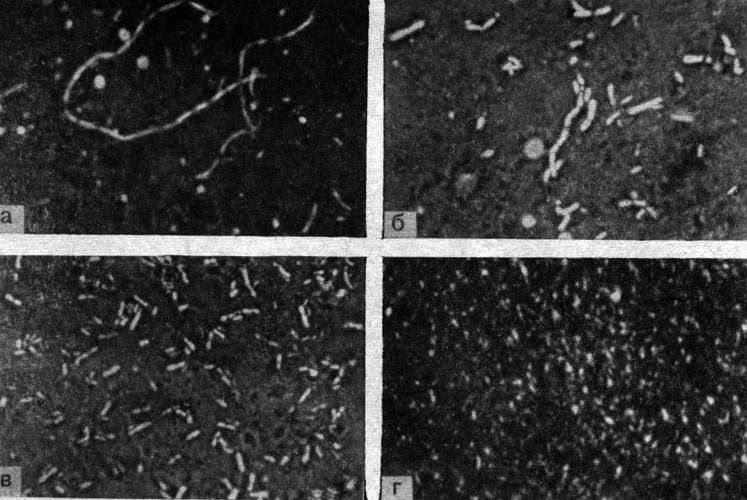
Рис. 2. Микрофото. Морфогенез риккетсий Провачека. Препараты из желточных оболочек куриных эмбрионов. Увеличено в 2000 раз. а - нитевидные формы (d), 3 - 5-й день после заражения; б - бациллярные формы (c), 5 - 6-й день после заражения; в - палочковидные формы (b), 6 - 7-й день после заражения; г - кокковидные формы (a), 7 - 9-й день после заражения. Оригинал
Указанная схема согласуется с представлениями П. Ф. Здродовского, Sikora и других авторов о характере и формах деления риккетсий. Очевидно, простейшими формами риккетсий являются однозернистые коккобациллы типа а. При определенных условиях среды эти формы воспроизводят поколение, состоящее из подобных им овоидов, в других же условиях размножающиеся коккобациллы не разделяются, как в предыдущем случае, ввиду чего их дочерние особи остаются связанными общей оболочкой, что приводит к возникновению палочковидных, бациллярных и нитевидных форм (би- , тетра- и полизернистые особи). Следовательно, полиморфизм возбудителя, по заключению П. Ф. Здродовского, морфологически можно определить как результат роста риккетсий с делением или без деления их на монозернистые, кокковидные особи.
Весьма интересную для понимания природы риккетсий проблему представляет вопрос о фильтрующихся формах риккетсий. Ранее дискутабельное представление о существовании фильтрующихся форм риккетсий нашло свое подтверждение после того, как для изучения морфологии риккетсий начали применять электронную микроскопию. В настоящее время убедительно доказано наличие фильтрующихся форм у риккетсий Бернета и Провачека (П. Ф. Здродовский, Е. М. Голиневич, 1956; Е. Я. Штернгольд, В. Н. Звагельская, 1952; В. И. Журбина, 1958; Kordova, 1959). Не исключается наличие их и у других видов риккетсий (у риккетсий Риккетса, Музера и возбудителя лихорадки цуцугамуси). В генетическом отношении фильтрующиеся формы, вероятно, соответствуют определенной фазе развития риккетсий, а именно являются ранней стадией внутриклеточной репродукции возбудителя.
Интересные сравнительноморфологические данные получены в результате сопоставления размера и формы различных видов риккетсий. Изложенные выше сведения относительно морфологии риккетсий основаны главным образом на изучении форм развития возбудителя эпидемического сыпного тифа, обладающего наиболее выраженным плеоморфизмом, на примере которого П. Ф. Здродовский (1948) предложил классификацию различных форм риккетсий (a, b, c и d).
Проведенные нами наблюдения за развитием риккетсий Провачека в желточных оболочках куриных эмбрионов позволили установить, что в течение первых 72 часов после заражения куриных эмбрионов риккетсии в клетках обнаружить не удается, тогда как спустя 5 - 8 дней отчетливо прослеживается процесс нарастания количества особей и уменьшения их размеров (см. рис. 2). В более ранних стадиях развития (4 - 6-й день) при скудном накоплении возбудителя в инфицированных клетках выявляются преимущественно палочковидные и нитевидные риккетсии. В поздних стадиях развития (7 - 12-й день) почти вся цитоплазма пораженных клеток заполнена мелкими кокковидными или коккобациллярпы - ми формами. Характерно, что в этой стадии роста очень редко видны крупные палочковидные формы и практически полностью отсутствуют нитевидные формы риккетсий. Это свидетельствует о том, что в процессе развития риккетсий происходит созревание возбудителя, выражающееся в массовом появлении зрелых кокковидных форм.
Наблюдая за развитием риккетсий Провачека в переживающих культурах тканей, Pinkerton и Hass еще в 1931 г. показали, что в пораженных клетках при скудном накоплении возбудителя, как правило, встречаются нитевидные формы длиной до 20 мк. Мелкие точечные формы обычно наблюдаются при обильном накоплении возбудителя в более поздних стадиях развития.
При выращивании риккетсий Провачека в тканевых культурах плеоморфизм был более выражен, чем в клетках инфицированных морских свинок и вшей. Одновременное присутствие в развивающейся культуре разных форм риккетсий может быть объяснено цикличностью развития возбудителя и накоплением разных фаз репродукции риккетсий в определенный промежуток времени.
А. В. Румянцев с соавторами (1947) пришли к выводу, что полиморфизм риккетсий, особенно характерный для роста в эпителии кишечника платяных вшей и реже в мезотелиальных и эпителиальных клетках легкого мышей и влагалищной оболочки яичка, объясняется наличием вегетативных форм (кокки и бациллы) хорошо адаптированного к выращиванию в соответствующих условиях штамма возбудителя. Другие нехарактерные для развития возбудителя формы (длинные нитевидные, гантелевидные), по мнению А. В. Румянцева, являются инволютивными или отмирающими формами. Эта точка зрения расходится с представлениями П. Ф. Здродовского и ряда других исследователей, а также противоречит нашим наблюдениям о нормальном физиологическом, а не патологическом характере плеоморфизма как естественного проявления динамики развития риккетсий и перехода их через различные стадии репродукции.
Риккетсии Провачека и риккетсии Музера в отличие от большинства других видов риккетсий, за исключением риккетсий Бернета, в инфицированных тканях могут полностью заполнять цитоплазму пораженных клеток, образуя своеобразные колонии - включения, получившие название "музеровских клеток".
Для риккетсий Музера весьма характерно интенсивное размножение в цитоплазме пораженных клеток мезотелия с обильным накоплением мельчайших кокковидных и палочковидных форм возбудителя. Практически недифференцируемые друг от друга мельчайшие формы риккетсий образуют сплошные скопления, целиком заполняющие цитоплазму пораженных клеток. Риккетсии Музера, давая весьма обильный рост в инфицированных клетках желточных оболочек куриных эмбрионов или мезотелия экспериментальных животных, обнаруживают менее выраженную склонность к плеоморфизму (реже образуют крупные палочковидные и нитевидные формы) и обладают меньшими размерами, чем риккетсии Провачека.
Для морфологии возбудителя Ку-лихорадки весьма характерны очень мелкие корпускулы, иногда на грани разрешающей способности оптического микроскопа. В этом случае на препарате из инфицированных желточных оболочек куриных эмбрионов видны мельчайшие пылевидные частицы преимущественно кокковидной или коккобациллярной формы. Крупные палочковидные и нитевидные формы встречаются сравнительно редко. В отличие от риккетсий сыпного тифа даже в ранних стадиях развития и при скудном накоплении преобладают формы a и b (рис. 3). Нитевидные формы не достигают таких больших размеров, как у риккетсий Провачека (10 - 40 мк), чаще состоят из 5 - 10 кокковидных особей и имеют более тонкую структуру. Располагаясь внутриклеточно, риккетсии Бернета редко заполняют целиком всю цитоплазму с образованием клеток музеровского типа даже при интенсивном инфицировании куриных эмбрионов или лабораторных животных. Своеобразие риккетсий Бернета заключается также в способности образовывать в пораженных клетках микроколонии (рис. 4), состоящие из большого числа особей, часто с вакуолизацией вокруг них протоплазмы, что особенно демонстративно при наблюдении за размножением возбудителя в культуре клеток (Smith, 1940; Freygang, 1949; Giroud, Gaillard, 1953; T. А. Бектемиров, 1955; В. А. Яблонская, 1956; И. В. Тарасович, 1957; О. С. Гудима, 1959).
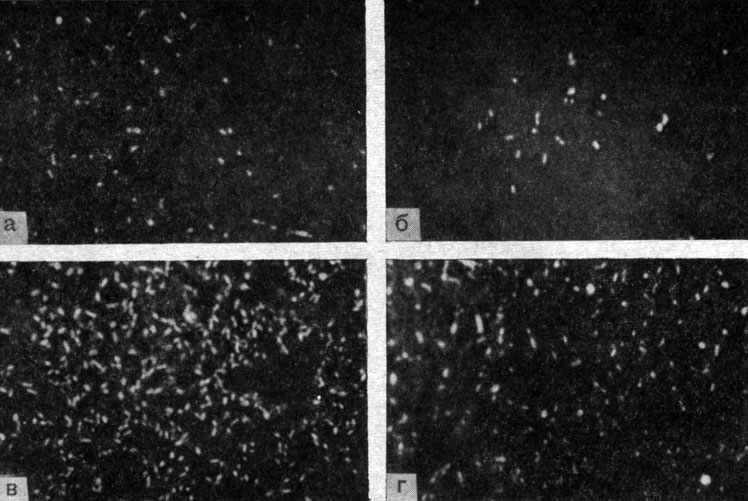
Рис. 3. Микрофото. Морфогенез риккетсий Бернета. Препараты из желточных оболочек куриных эмбрионов. Увеличено в 2000 раз. а - 4 - 5-й день после заражения, накопление на 1 крест; б - 5 - 6-й день после заражения, накопление на 2 креста; в - 6 - 7-й день после заражения, накопление на 3 креста;: г - 7 - 9-й день после заражения, накопление на 4 креста. Оригинал
Величина отдельных частиц возбудителя Ку - лихорадки варьирует от невидимых в световом микроскопе фильтрующихся форм (Сох, Bell, 1939) до крупных риккетсий, напоминающих мелкие бактерии. Размеры риккетсий Бернета колеблются от 0,25×0,25 до 0,25×1,5 мк (Сох, 1938; Herzberg с соавторами, 1950; С. М. Кулагин и Н. Г. Кекчеева, 1954; А. Ю. Иллютович, 1957). По данным электронной микроскопии, величина палочковидных форм риккетсий Бернета колеблется от 0,2 до 0,4 мк в ширину и от 0,3 до 0,9 мк в длину (Wiesmann, 1948; Van Rooyen, Scott, 1949; Liebermeister, Zehender, 1950). При изучении ультратонких срезов возбудителя было установлено, что диаметр риккетсий не превышает 0,2 мк (Stoker и соавторы, 1956).
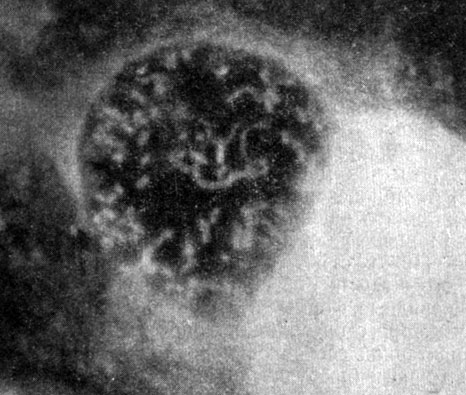
Рис. 4. Микрофото. Микроколония риккетсий Бернета внутри вакуоли в клетках L. Люминесцентная микроскопия. Увеличено в 1800 раз. Оригинал
Сравнительная оценка морфологии австралийских, американских, западноевропейских и отечественных штаммов риккетсий Бернета показала, что риккетсии разных штаммов могут отличаться по своей величине. Так, В. А. Яблонская (1956) и Т. А. Бектемиров (1956) установили, что риккетсии крымского штамма № 73 несколько меньше ряда других штаммов, a Hengel с соавторами (1960) отметили различие между риккетсиями немецкого штамма Zuzenhaus, имеющего преимущественно кокковидные формы, и американского штамма Nine Mile, у которого преобладают палочковидные формы.
Характерной особенностью обладают риккетсии клещевой группы (возбудители пятнистой лихорадки Скалистых гор, марсельской лихорадки, клещевого сыпного тифа Северной Азии и др.), которые, являясь типичными внутриклеточными паразитами, в отличие от других видов риккетсий способны поражать не только цитоплазму, но и ядра инфицированных клеток (Wolbach, 1919; Pinkerton, Hass, 1932, 1936). Типичным представителем этой группы риккетсий, объединенных под названием Dermacentroxenus, является возбудитель пятнистой лихорадки Скалистых гор, открытый и впервые изученный Ricketts (1906, 1909), показавшим наличие в крови больных людей, морских свинок и обезьян, а также в искусственно инфицированных на больных животных клещах, мелких телец, напоминающих по форме диплококки, разъединенных небольшим количеством промежуточной субстанции. Обстоятельные исследования Wolbach (1918, 1919) позволили выявить три основные морфологические формы возбудителя пятнистой лихорадки Скалистых гор - риккетсий Риккетса. Первый тип, по Wolbach, представляет собой относительно крупные, расположенные в ядре клеток, бациллоподобные тельца, лишенные хроматиновых гранул. Второй тип - мелкие палочковидные тельца с хроматиновыми гранулами. Третий тип - относительно крупные ланцетовидные парные тельца. По данным Wolbach, эти тельца часто окружены очень нежной, но достаточно четкой, неокрашиваемой капсулой, толщина стенки которой на приведенных автором рисунках достигает иногда диаметра самих телец. Как свидетельствуют наблюдения Wolbach, наиболее характерны тельца второго типа, представляющие стадию "покоя", тогда как два других типа возбудителя являются стадиями "размножения".
По данным Nicholson (1923), в ранний период заболевания в органах морской свинки содержались диплобациллярные, а в более поздний срок - бациллярные формы риккетсий, однако описанных Wolbach кокковидных и диплококковых форм автор не обнаружил.
Spenser и Parker (1926, 1930) отметили, что во взрослых накормленных иксодовых клещах, ранее в фазе личинки инфицированных риккетсиями Риккетса, обычно содержатся мелкие кокковидные, ланцетовидные и диплобациллярные тельца. В аналогичных партиях клещей, но находившихся в течение нескольких недель при комнатной температуре, риккетсии были крупнее. Кроме того, встречались нитевидные формы. В ряде случаев в клещах, в которых титрованием на морских свинках определялась высокая концентрация риккетсий, микроскопически обнаруживалось небольшое накопление их, однако часто в значительном количестве встречались гранулы, которые, по мнению Spenser и Parker, могли являться прериккетсиозной стадией развития риккетсий Риккетса. Природа этих форм риккетсий была выяснена исследованиями Plotz с соавторами (1943). Оказалось, что в ряде случаев в пораженных клетках содержатся мелкие кокковидные риккетсии, которые легко обнаруживаются под электронным микроскопом, но с большим трудом дифференцируются в обычном микроскопе от элементов ткани.
Значительный полиморфизм риккетсий Риккетса был обнаружен при культивировании в переживающей ткани и при пассажах через легочную ткань белых мышей. Так, по данным Giroud и Panthier (1941), в легких мышей, зараженных через нос под эфирным наркозом, встречаются мелкие точечные и крупнее фрагментированные тельца, причем длина последних достигает 3 - 4 мк. В переживающей ткани в опытах Pinkerton (1934, 1942) риккетсии Риккетса встречалась в форме крупных ланцетовидных телец, иногда достигавших размера пневмококка, однако в ядрах тех же клеток имелись скопления мелких парных телец. В цитоплазме клеток обнаруживались мелкие и более крупные диплобациллярные тельца.
Как свидетельствуют наблюдения Schaechter, Bozeman и Smadel (1957), в живых клетках фибробластов крыс (штамм 14pf) риккетсии Риккетса встречаются по всей цитоплазме и иногда в ядрах. Риккетсии часто обнаруживаются в межклеточном веществе, что в большинстве случаев обусловлено разрушением пораженных клеток, которое обычно происходит на 5 - 6-й день после заражения культуры клеток, несмотря на относительно необильное накопление риккетсий к этому сроку.
Значительный интерес представляют наблюдения названных авторов в отношении механизма выхода риккетсий Риккетса из живых клеток фибробластов крыс 14pf. Ими было показано, что в ряде случаев риккетсии располагаются в той области цитоплазмы клетки, в которой имеются микрофибриллы, причем риккетсии часто оказываются соединенными с сокращающейся частью клетки. При сокращении микрофибриллы риккетсии либо увлекаются в цитоплазму, либо выталкиваются из клетки и оказываются в межклеточном веществе. Таким образом, внеклеточное расположение риккетсий Риккетса обусловлено не только разрушением клетки, но и выходом риккетсий из живых клеток. Schaechter, Bozeman и Smadel удалось наблюдать поперечное деление палочковидных форм риккетсий Риккетса с образованием диплобациллярных телец. Процесс деления риккетсий в условиях тканевой культуры фибробластов крысы 14pf занимал в среднем 120 минут. Образовавшиеся диплобациллярные тельца оставались неизмененными в среднем до 159-й минуты, после чего два тельца диплобациллы разъединялись и расходились друг от друга с образованием двух самостоятельных телец.
Таким образом, в указанных исследованиях, проведенных с живыми риккетсиями Риккетса, было установлено, что диплобациллярные тельца представляют раннюю стадию поперечного деления риккетсий, небольшие кокковидные и бациллообразные тельца - риккетсий вскоре после деления, а более крупные бациллярные тельца - "зрелые" формы риккетсий. Указанные формы риккетсий Риккетса, наблюдавшйеся в условиях живой ткани, в основном совпадают с морфологическими разновидностями возбудителя пятнистой лихорадки Скалистых гор, описанными Ricketts, Wolbach, Nicholson, Parker, Spenser, Pinkerton и другими исследователями в фиксированных препаратах, приготовленных из инфицированных тканей животных, людей и клещей. Следовательно, различные морфологические формы возбудителя пятнистой лихорадки Скалистых гор оказались этапами онтогенетического развития риккетсий Риккетса, а одновременное наличие форм, свойственных различным этапам развития риккетсий, обусловило их полиморфизм, отмеченный многими исследователями.
Существенной особенностью риккетсий Риккетса является способность к размножению внутри ядер пораженных клеток. Wolbach, впервые выявивший внутриядерные формы риккетсий Риккетса, обнаружил их только в некоторых клетках инфицированных нимф и имаго клеща D. andersoni (Wolbach, 1918, 1919, 1921, 1925). В последующем внутриядерное размножение риккетсий Риккетса в клетках клещей было подтверждено рядом исследователей. Внутриядерные формы риккетсий были обнаружены Pinkerton (1934) в клетках переживающей ткани оболочки яичка морской свинки, a Schaechter, Bozeman, Smadel (1957) - в живой культуре фибробластов крысы 14pf. В тканях млекопитающих, несмотря на многочисленные эксперименты, долгое время не удавалось выявить риккетсии Риккетса внутри ядер клеток, что позволило Pinkerton (1934) высказать предположение об отсутствии таких форм в клетках животных. Однако позднее внутриядерное расположение риккетсий Риккетса удалось установить при окраске парафиновых срезов из яичка и влагалищной оболочки яичка больных морских свинок (Pinkerton, 1948). Используя фиксацию в жидкости Pyro и окраску парафиновых срезов по методу Романовского, Pinkerton (1948) выявил внутриядерное расположение риккетсий Риккетса также в клетках желточной оболочки инфицированных куриных эмбрионов.
В клещах, по наблюдение Wolbach (1919), внутриядерные формы риккетсий Риккетса встречаются в клетках мальпигиевых органов, ректального мешка и слюнных желез. В нимфах риккетсии в значительном количестве были обнаружены в ядрах гиподермы, а в имаго - в клетках мускулатуры. По мнению Wolbach, для риккетсий Риккетса, расположенных внутри ядер пораженных клеток, характерны мелкие формы, причем наличие таких телец вне ядер обусловлено ранением клеток в процессе изготовления препаратов. По данным Pinkerton (1934, 1942), в переживающей ткани влагалищной оболочки яичка морской свинки внутриядерные формы риккетсий Риккетса представляют компактные скопления, окруженные четкой просветленной зоной, а иногда мелкие парные гранулы, причем в цитоплазме тех же клеток имеются не только мелкие, но и более крупные диплобациллярные и ланцетовидные тельца. Ядра, в которых имеются скопления риккетсий, значительно увеличены.
В культуре клеток фибробластов крысы 14pf риккетсии Риккетса обычно встречаются в виде отдельных телец, лишь изредка расположенных внутри ядер, причем закономерного поражения клеток не отмечается (Scha - echter с соавторами, 1957).
Таким образом, внутриядерные формы риккетсий Риккетса наиболее постоянно встречаются в клетках инфицированных клещей D. andersoni и в тканевых культурах.
Способность риккетсий Риккетса к размножению внутри ядер пораженных клеток явилась важным признаком при дифференциации их от ряда других видов риккетсий, что и было учтено Pinkerton (1936) при классификации риккетсий для выделения рода Dermacentro - xenus.
Наиболее типичные для риккетсий Риккетса палочковидные формы, по данным обычной микроскопии, достигают в длину 1 мк, а в ширину 0,3 - 0,2 мк (Becker, 1926). Длина бразильского варианта риккетсий Риккетса варьировала от 1,4 до 0,5 мк (Monteiro, 1931). В клещах D. andersoni, по данным электронной микроскопии, размер риккетсий Риккетса достигает в длину 1,2 мк, а в ширину 0,6 мк (Van Rooyen, Scott, 1949). Длина риккетсий в культуре клеток фибробластов крысы 14pf составляла 1,5±0,4 мк, а, ширина 0,55±0,06 мк (Schaechter с соавторами, 1957). Наконец, ланцетовидные формы риккетсий Риккетса (третий тип по Wolbach) достигают в длину 1 мк, а в ширину 0,3 - 0,2 мк (Wolbach, 1919).
Наиболее типичными морфологическими формами другого представителя группы Dermacentroxenus - возбудителя марсельской лихорадки (риккетсий Конора) - у морских свинок и в тканевых культурах являются палочковидные и более крупные бациллярные особи. Кроме того, изредка обнаруживаются гигантские нитевидные образования варьирующей конфигурации. В отличие от риккетсий Провачека риккетсии Конора характеризуются более грубой формой и весьма скудным накоплением в цитоплазме. Возбудитель марсельской лихорадки обычно диффузно расположен в цитоплазме и никогда не заполняет клетки целиком, что характерно для возбудителя сыпного тифа. Внутриядерное размножение характерно как для средиземноморских, так и для черноморских штаммов риккетсий Конора, выделенных в 1947 - 1948 гг. П. Ф. Здродовским и Е. М. Голиневич. Внутриядерное расположение риккетсий прекрасно выявляется в клетках мезотелия влагалищной оболочки тестикул морских свинок и желточных оболочек куриных эмбрионов при окраске препаратов карболфуксином и метиленовой синью (по методу П. Ф. Здродовского). В этом случае внутри ядер пораженных клеток контрастно выделяются рубиново - красные риккетсии на фоне окрашенного в синий цвет ядерного вещества.
Подробно изученный советскими исследователями (Е. И. Милль, 1936; Н. И. Антонов, А. Г. Найштат, 1936, 1937; М. К. Кронтовская, М. Д. Шматиков, 1943; С. М. Кулагин, 1945, 1953; С. П. Пионтковская, О. С. Коршунова, 1940; А. А. Преображенский, 1946; Н. В. Вощакина, 1948; Е. М. Голиневич, 1949, 1952) возбудитель клещевого сыпного тифа Северной Азии на основании цитологических и серологических свойств был отнесен к роду Dermacentroxenus (Е. М. Голиневич, 1949). По своей морфологии данный возбудитель близок к риккетсиям марсельской лихорадки. Для возбудителя клещевого сыпного тифа Северной Азии характерно наличие палочковидных и бациллярных форм при относительно редком обнаружении нитевидных грубых образований, а также умеренном или скудном заполнении цитоплазмы пораженных клеток и отсутствии, даже в случаях заражения морских свинок большими дозами возбудителя, скоплений риккетсий, напоминающих музеровские клетки. Однако С. М. Кулагин (1954) показал, что при внутри - брюшинном заражении хлопковых крыс смертельными дозами возбудителя в пораженных клетках мезотелия брюшины встречаются клетки типа музеровских. Кроме того, колонии риккетсий можно обнаружить при периор - хите у зараженных морских свинок в ядрах клеток мезотелия при окраске препаратов карболфуксином и метиленовой синью (П. Ф. Здродовский, Е. М. Голиневич, 1956).
С точки зрения сравнительной оценки размеров риккетсий значительный интерес представляют исследования И. Н. Кокорина и Н. Н. Рыбкиной (1960), которые применили методику оценки величины риккетсий по оптическому бактериальному стандарту в сравнении с размером брюшнотифозных бактерий. Авторы установили, что в расчете на 1 млрд. брюшнотифозных палочек в 1 мл очищенной риккетсиозной суспензии содержится 1,53 млрд. особей риккетсий Провачека, 4,77 млрд. риккетсий Музера и 6,5 млрд. риккетсий Бернета. Очевидно, что размер частиц риккетсий Провачека больше, чем у других риккетсий, и приближается к размеру брюшнотифозных палочек (Salmonella typhi), которые имеют величину порядка 0,5 - 0,8×1,5 - 3 мк (М. Н. Лебедева, 1960). Риккетсии Музера меньше риккетсий Провачека примерно в 3 раза, а риккетсии Бернета - в 4 раза. Полученные расчетные данные соответствуют фактическим размерам указанных риккетсий, определенным другими методами (центрифугирование, фильтрация, электронная микроскопия).
Изучение размеров риккетсий в очищенных препаратах при электронной микроскопии позволило уточнить некоторые величины, полученные при ультрафильтрации. Так, Van Rooyen и Scott (1949) при электронной микроскопии определили, что размеры риккетсий Бернета колеблются от 0,25×0,5 до 0,25×1,5 мк. О. С. Гудима (1959, 1961), изучая динамику роста риккетсий Бернета в культуре клеток HeLa, установил, что частицы этого возбудителя имеют форму кокков и палочек и размеры 0,3 - 0,4×0,3 - 0,7 мк, а в культурах, содержавших 100 ЕД пенициллина, приобретают форму крупных (0,7 - 1,1×1,5 - 2,2 мк), округлых или овоидных образований. Bobb и Downs (1962) наблюдали в электронном микроскопе разные фазы развития риккетсий Бернета. После очистки риккетсий дифференциальным центрифугированием авторы готовили препараты риккетсий, напыленных хромом. На электрономикрограммах наряду с кокковидными формами риккетсий (размером 0,3×0,5 мк) встречались спиралеподобные структуры величиной 0,5×0,3 мк. Размеры и общий вид отдельных указанных форм позволяют полагать, что это не простые риккетсиозные элементы, а сложная структура полярно расположенных элементарных частиц риккетсий, заключенных в общую оболочку.
Plotz с соавторами (1943), изучая морфологическую структуру риккетсий, высказали предположение, что риккетсии по своему строению занимают промежуточное положение между фильтрующимися вирусами и бактериями. При электронной микроскопии было установлено структурное сходство риккетсий Провачека, Музера, Бернета и Риккетса, но различие в размерах, так как риккетсии Бернета оказались почти в 2 раза меньше других риккетсий. Наличие у риккетсий четкой мембраны, окружающей цитоплазму, в которой находятся гранулы, делает это морфологическое сходство с бактериями более полным. Plotz удалось проследить стадии бинарного деления риккетсий, когда коккобацилла приобретает гантелевидную форму, в которой цитоплазма постепенно делится на две части, временно объединенные общей оболочкой, которая в конечном итоге раздваивается, в результате чего образуются две дочерние риккетсиозные частицы. Подобная же картина отмечалась нами при изучении возбудителей клещевых риккетсиозов, сыпного тифа и Ку-риккетсиоза (рис. 5). Аналогичные результаты были получены Weiss (1943) при электронной микроскопии риккетсий Провачека. Автор наблюдал процесс деления бациллярных форм с образованием кокковидных. Размер кокковидных и коккобациллярных форм колебался в пределах 0,5×0,5-0,5×1-1,5 мк, т. е. несколько превышал размеры риккетсий Бернета, приведенные Bobb и Downs.
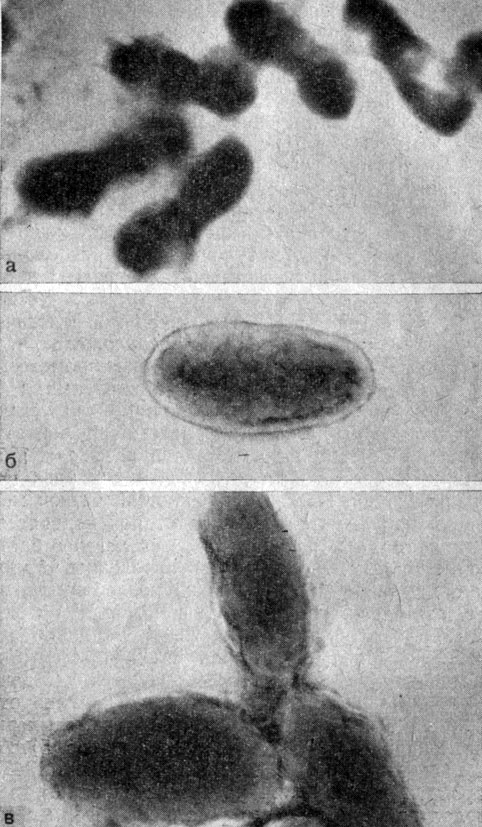
Рис. 5. Микрофото. Электронная микроскопия. Увеличено в 20 000 раз. а - риккетсии Провачека; б, а - риккетсии Бериета. Оригинал
Наибольший интерес представляет изучение при электронной микроскопии тонкой структуры риккетсий с применением методики ультратонких срезов зараженных клеток, заключенных в метилметакрилат. Rosenberg и Kordova (1960) через 48 часов после заражения в переживающих кусочках желточной оболочки в растворе Гироде наблюдали образования, представляющие скопления мелкогранулированных гомогенных частиц (матрицы), лишенных наружной мембраны, или компактных, непроницаемых для электронов масс вещества. Через 4 - 6 дней отмечали наличие риккетсий Бернета размером 0,3 - 1,2 мк. Риккетсии имели компактную тонкогранулированную субстанцию, напоминающую по структуре вироплазму. Частицы содержали наружную мембрану и плотное центральное тело, непрозрачное для электронов. Более крупные формы риккетсий (2 мк) вместо плотного центрального тела имели небольшие гранулы, диффузно расположенные в вироплазме. Кроме того, встречались промежуточные формы в состоянии сегментации. В этом случае наблюдались двойные компактные структуры, окруженные общей мембраной и имеющие тонкую перегородку.
Anacker, Fukushi, Pickens, Lackman (1964), наблюдая за динамикой развития риккетсий Бернета при электронной микроскопии препаратов желточной оболочки куриных эмбрионов, установили наличие эклипс - фазы и показали, что для ультратонкой структуры возбудителя; характерно наличие плотного фибриллярного нуклеоида, окруженного менее плотной цитоплазмой, содержащей гранулы диаметром около 15 ммк.
При электронномикроскопическом изучении структуры риккетсий Провачека в ультратонких срезах клеток желточных оболочек куриных эмбрионов Г. П. Гандзий, Б. Г. Затуловский, Л. Я. Школьник и Н. М. Шестопалова (1964, 1965, 1966) обнаружили четко выраженную многослойную оболочку, состоящую из наружной осмиофильной (2 слоя по 25 Å), средней и внутренней осмиофобной мембран толщиной ≈30Å. В неоднородной по плотности цитоплазме авторы отметили наличие канальцев, скопление осмиофильного вещества на полюсах риккетсий, поперечных мембран и двух видов гранул. Оптически более плотные гранулы размером от 70 до 700Å напоминают по своей структуре рибосомы. Второй тип гранул, размером значительно превышающих рибосомы, представляет скопление оптически менее плотного вещества.
Ito и Winson (1965) при электронномикроскопическом изучении тонкой структуры возбудителя волынской лихорадки, культивируемого в культуре клеток и в организме вшей, определили размер, равный 0,2 - 0,5×1,6 мк, около 80Å и плазматическую мембрану ≈70Å. В цитоплазме обнаружены рибосомы до 150Å в диаметре, РНК и ДНК. Различий в структуре риккетсий, выращенных в клетках и во вшах, не установлено. Показано, что ядерный материал диффузно расположен в цитоплазме. Наличие клеточной мембраны у риккетсий под электронным микроскопом отметил Fuller (1959), показавший также содержание в вироплазме риккетсий ДНК и РНК. Allison и Perkins (1960) на ультратонких срезах под электронным микроскопом выявили у риккетсий наличие двойной мембраны. Эти данные согласуются с нашими исследованиями тонкой структуры риккетсий Провачека и риккетсий Бернета на ультратонких препаратах под электронным микроскопом. На электрономикрограмме (рис. 6) среза риккетсий Провачека из очищенной культуры желточных оболочек куриных эмбрионов отчетливо видна двойная мембрана, ограничивающая зернистую вироплазму, в которой выделяется оптически более плотное образование - нуклеоид. Нуклеоид имеет спиралевидную или гранулярную структуру. Можно полагать, что спиралевидные и диффузно расположенные гранулы представляют скопление нуклеиновых кислот. Аналогичную структуру имеют на ультратонких срезах и риккетсии Бернета (рис. 7).
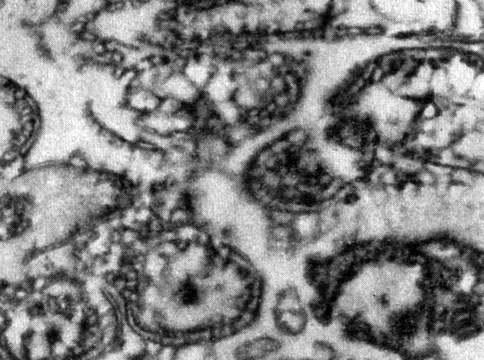
Рис. 6. Микрофото. Ультратонкая структура риккетсий Провачека. Электронная микроскопия. Увеличено в 20 000 раз. Оригинал
Морфологические формы риккетсий, представленные на элекгронограммах, позволяют проследить процесс деления риккетсий на разных стадиях развития. Так, имеются особи с наметившейся гантелевидной перетяжкой и частица, нуклеоид которой уже разделен на две части, но полного деления клетки, объединенной общей оболочкой, еще не произошло. Имеются также риккетсии, завершившие деление с образованием дочерних особей, содержащие нуклеоид, вироплазму и окруженные оболочкой.
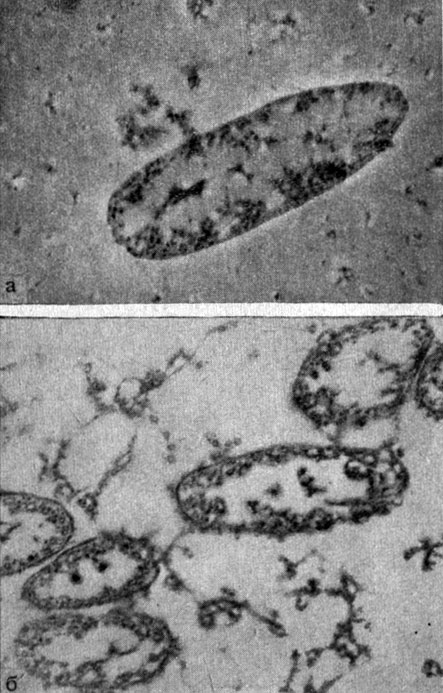
Рис. 7 (а, б). Микрофото. Ультратонкая структура риккетсий Бернета. Электронная микроскопия ультратонкого среза риккетсий в культуре клегок. Увеличено в 20 000 раз. Оригинал
В заключение следует отметить, что ряд исследователей пытался установить зависимость между морфологией риккетсий и их вирулентностью. Так, Kausche и Sheiris (1951) считают, что культуры, состоящие из кокковидных форм, отличаются высокой вирулентностью, тогда как наличие полиморфных форм с преобладанием палочковидных риккетсий часто указывает на малую вирулентность, что, по мнению авторов, можно наблюдать у штамма Nine Mile риккетсий Бернета. По мнению Ш. Д. Мошковского (1945), нормальными жизнеспособными формами риккетсий являются кокковидные и коккооациллярные, тогда как палочковидные и нитеподобные образования представляют скопления риккетсий, окруженных капсулой и, следовательно, находящихся в состоянии анабиоза по аналогии с капсулообразующими бактериями. С другой стороны, в соответствии с данными П. Ф. Здродовского и Е. М. Голиневич (1956) все формы жизнеспособных риккетсий, обладают одинаковой вирулентностью, так как они являются нормальными формами развития возбудителя.
В последние годы получены убедительные данные, свидетельствующие о существенных изменениях вирулентности риккетсий без изменения их морфологии (физиологическая изменчивость) и о наличии одинаковой морфологии у штаммов, резко отличающихся по вирулентности (см. ниже).
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'