1.2. Популяция под действием вектора отбора, идущего по пластичным признакам фенотипа
Все предыдущее изложение вопросов направленности микроэволюции велось при трех существенных упрощениях: 1) признаки, по которым идет отбор, предполагаются стабильными, лишенными изменений в пределах нормы реакции,, точнее, не имеющими таковой; 2) на признак действует лишь один вектор отбора; 3) признаки не взаимодействуют между собой. Между тем пространство адаптации популяции многомерно. В каждый момент оно определяется всеми параметрами экологической ситуации, действующей на всю организацию особи, т. е. на все ее прямо или опосредованно взаимодействующие со средой признаки, которые в свою очередь прямо или косвенно связаны между собой (организм целостен), а также на связи между особями данной популяции.
Большая часть условий внешней среды, в которых существуют популяции любого дикого вида, испытывает колебания, причем эти колебания по многим параметрам не закономерны и во времени и в пространстве. Это относится к абиотическим (микроклимату, химизму среды и т. п.) и в большей степени к биотическим факторам. Поэтому особи любого вида вынуждены адаптироваться не к конкретным значениям того или иного внешнего воздействия, а к. некоторому диапазону его изменений. Это общеизвестное положение применимо не только к отдельной особи, но и к популяции в целом. В зависимости от типа онтогенеза, с которым связан характер генетического определения признаков (Шмальгаузен, 1968 а), можно выделить два полярных способа адаптации популяций к незакономерным изменениям среды: полиморфизм генотипически детерминированных признаков (Гриценко и др., 1983; Левонтин, 1978) и модификационный полиморфизм (Шмальгаузен, 1968 а, 1982). Оба способа адаптации связаны непрерывным рядом переходов, причем промежуточными членами этого ряда являются полигенные признаки и эпигеномные со стабильным фенотипическим выражением. Различие с точки зрения адаптации между крайними членами ряда заключается преимущественно в том, что в случае сбалансированного полиморфизма генофонд популяции подстраивается к изменениям среды как целое, платя за адаптацию гибелью особей, обладающих пониженной в данных условиях жизнеспособностью. При адаптации через изменения в пределах нормы реакции приспособление достигается за счет онтогенетических, физиологических и поведенческих реакций индивидуумов, причем плата за адаптацию выражается в гибели (или устранении от размножения) особей, недостаточно толерантных к данному воздействию. Согласно И. И. Шмальгаузену (1968а), в этих условиях плата за эволюцию может быть существенно меньшей, а адаптация более быстрой. Поскольку возникновение каждого из указанных способов обусловлено многими причинами, а большая или меньшая их адаптивность определяется конкретной экологической ситуацией, оценить, какой из способов адаптации лучше, не представляется возможным. Надо, однако, отметить, что большая часть приспособлений любых эукариотических организмов к меняющимся условиям среды достигается за счет онтогенетических, физиологических и поведенческих реакций, т. е. пластичных (лабильных, по терминологии Шмальгаузена, 1968а, 1982) признаков фенотипа, обладающих более или менее широкой нормой реакции.
Если не считать Ламарка и ламаркистов, то первым, кто обратил внимание на индивидуальную адаптацию организмов в процессе эволюции, был, вероятно, Болдуин (цит. по Майр, 1968), введший принцип генокопирования. Основной вклад в изучение механизмов эволюции подобных признаков внесли И. И. Шмальгаузен (1968а, 1982) и С. Уоддингтон (Waddington, 1957). С. Уоддингтону принадлежит экспериментальное доказательство генокопирования. При отборе по температурному морфозу - разрыву поперечной жилки крыла у дрозофилы (фенотип Crossveinless) было получено увеличение частоты его возникновения с повышением температуры и появление этого фенотипа в случае нормальной температуры.
И. И. Шмальгаузен разработал теорию формирования адаптивных модификаций и эволюции форм с пластичными признаками фенотипа. При этом он рассматривал три основных случая направленной эволюции организмов, способных к адаптивным модификациям: лабильный организм в однородно меняющейся среде, лабильный организм в разнородно меняющейся среде и стабилизацию новой адаптивной нормы. Эволюция пластичных признаков фенотипа в однородно меняющейся среде при движущей форме отбора (первый случай) отличается от эволюции стабильных признаков тем, что элиминируются не только неблагоприятные наследственные уклонения, но и те адаптивные модификации, которые возникли в ответ на случайные изменения внешней среды и стали не адаптивными при ее возвращении к норме. В то же время выживают не только особи, наследственные уклонения которых совпадают с изменением среды, но и все особи, проявления нормы реакции которых соответствуют этим изменениям, т. е. вьь живание обусловлено и генотипической и паратипической изменчивостью. В результате сочетания наследственных и модификационных изменений происходит генокопирование, и те признаки, которые были первоначально лишь вариантами фенотипа в пределах нормы реакции, фиксируются генетически как единственное проявление данного признака. Так, рассеченность листьев у Аnеmоnае pulsatila в западной части ареала сильнее, чем в восточной, а цветки поникшие, причем при скрещивании западных и восточных форм признаки менделируют. Однако обе формы листьев можно получить у одного растения, меняя степень освещенности, т. е. модификационно. При более сухом климате Восточной Европы освещенность выше и модификация фиксированна (Шмальгаузен, 1968а).
В разнородно меняющейся среде, согласно И. И. Шмальгаузену, отбор действует на пластичные признаки фенотипа, создавая систему более или менее дискретных проявлений нормы реакции. Примерами служат три модификации листьев стрелолиста: лентовидные - подводные, округлые - плавающие и стреловидные - воздушные. И. И. Шмальгаузен подчеркивал, что проявления подобных модификаций стабилизируются в ходе эволюции за счет совершенствования морфогенетических механизмов, происходящего посредством элиминации случайных уклонений от оптимального пути. Зависимые от среды пути развития приобретают при этом авторегуляторный характер. Так, например, световые или теневые листья развиваются в зависимости от освещенности почек в предыдущем сезоне вегетации.
И. И. Шмальгаузену же принадлежит наиболее ясное обоснование значения адаптивных модификаций, как фактора, определяющего быструю адаптацию популяций к новым условиям среды. При изменении внешних условий, если диапазон этих изменений не выходит за пределы нормы реакции особей данной популяции, особи, ее составляющие, за одно поколение подстраиваются к изменению. Элиминация при таком способе адаптации снижается по сравнению с адаптацией стабильных организмов. Затем постепенно, под действием стабилизирующего отбора, фиксируется оптимальная для новых условий новая норма реакции.
Таким образом, учет наличия пластичных признаков фенотипа, не меняя основной концепции - направленного комбинирования генов под действием отбора, дает возможность объяснить ускорение темпов адаптации при изменениях внешних условий, в то же время подчеркивает значение среды в определении направления эволюции. Действительно, если при отборе по стабильным признакам направление отбора зависит и от среды, и от наличной генотипической изменчивости, способной подвергаться отбору в данном направлении, то при эволюции признаков с широкой нормой реакции адаптация происходит независимо от наследственной изменчивости. Отбор и комбинирование генов отстают по времени от адаптации и не определяют ее направление.
Анализ эволюции пластичных признаков фенотипа ставит вопрос о том, как возникают подобные признаки. И. И. Шмальгаузен был, по-видимому, первым, кто пытался решить этот вопрос. На основе развитого им представления об эволюции процессов онтогенеза под действием стабилизирующего отбора он предположил, что в колеблющихся условиях среды особи, находящиеся на краях распределения по отбираемому признаку, в зоне условной элиминации, и обладающие генотипически обусловленной более широкой нормой реакции, получают селективное преимущество, что ведет в ряду поколений к формированию адаптивных модификаций. Это положение сводится, по сути дела, к следующему дарвиновскому тезису: чтобы вести отбор в каком-либо направлении, должна существовать изменчивость, которая может быть подвергнута отбору в данном направлении. В приложении к рассматриваемому случаю это означает, что для начала отбора на расширение нормы реакции должна существовать изменчивость по широте нормы реакции отбираемых признаков.
Подобная изменчивость, по-видимому, чаще возникает, когда мутации меняют эпигенетические механизмы онтогенеза, дестабилизируя их. Например, в норме и окраска глаз, и морфология их у дрозофилы - признаки стабильные. Мутации, приводящие к изменению окраски глаз, такие как cherry, eosin, apriootte, white и т. п., нарушая биосинтез пигментов глаза, т. е. действуя на биохимическом уровне, не обладают сколько-нибудь заметной широтой нормы реакции. Напротив, мутации, меняющие число или строение фасеток глаза (eyeless, bar, ultrabar, puffed и др.), т. е. мутации, действующие на морфогенез, всегда обладают более или менее широкой нормой реакции (Митрофанов, 1978; Тимофеев-Ресовский, Иванов, 1966). Изменчивость по широте нормы реакции в природных популяциях дрозофилы показана Н. В. Глотовым и В. В. Таракановым (1985). Если вышеизложенное справедливо, то вопрос о первоначальном (в филогенетическом смысле) возникновении признаков с широкой нормой реакции сводится к вопросу о возникновении эпигенотипа, т. е. выходит за рамки задач данной работы. Так или иначе мутации, меняющие широту нормы реакции, существуют, и, следовательно, материал для отбора в данном направлении имеется. Индивидуальная изменчивость пластичных признаков фенотипа, в первую очередь физиологических реакций, начинает привлекать внимание биологов. Ряд примеров приводит А. М. Уголев (1985), много примеров содержится в работах энтомологов (Семевский, Семенов, 1982; Schwerdtfeger, 1963). Индивидуальная изменчивость пластичности поведенческих признаков общеизвестна.
Однако в других отношениях предложенное И. И. Шмальгаузеном объяснение расширения нормы реакции не может быть принято по следующим причинам:
1. Согласно этой модели стабилизирующий отбор ответствен не только за расширение, но и за сужение нормы реакции - генотипическое фиксирование модификаций, которые происходят через элиминацию неадаптивных к данным условиям ее проявлений. Если учесть, что внешние условия флуктуируют постоянно, становится непонятным, когда один и тот же селективный механизм ведет к сужению, а когда - к расширению нормы реакции.
2. Стабилизирующий отбор действует на края распределения, а центральная его часть - популяционная норма - отбору не подвергается по определению. Следовательно, расширение нормы реакции у особей, относящихся к центральным классам распределения, должно происходить через их скрещивание с особями, получившими селективное преимущество на его краях. Это привело бы oк тому, что в череде поколений расширение нормы реакции у особей центральных классов распределения отставало бы от такового у боковых классов. Тем самым особи из центра распределения должны в каждый данный момент времени иметь более узкую норму реакции, чем особи, уклоняющиеся от средней. В действительности этого не наблюдается (см. ниже). В литературе также не удалось найти подобных указаний.
3. Колебания внешних условий для особей, вынужденных к ним приспосабливаться, представляют собой попеременные ухудшения и улучшения экологической обстановки. В первом случае селективное преимущество будут получать особи, близкие к модальному классу, и особи, способные приблизиться к нему благодаря широте их нормы реакции. Однако при улучшении условий особи, обладающие более широкой нормой реакции, не получат селективного преимущества, так как в этой ситуации будут выживать организмы с любой нормой реакции. Следовательно, стабилизирующий отбор всегда способствует сужению нормы реакции (фиксации модификаций), но не ее расширению.
Исходя из изложенных соображений, нами была поставлена серия опытов на головастиках Rana temporaria и R. esculenta с целью выяснить механизм расширения нормы реакции (Северцов, 19816, 1984; Северцов, Сурова, 1981; Сурова, 1985). В качестве примера пластичного признака фенотипа были использованы темпы роста головастиков. Среди факторов, модифицирующих скорость роста личинок бесхвостых амфибий, таких как температурный режим, газовый режим, освещенность, обеспеченность пищей и т. п., важное значение имеют внутрипопуляционные взаимодействия развивающихся особей. С повышением плотности головастиков (выше трех особей на 2 л воды) средние темпы роста и развития снижаются, а изменчивость по этому признаку растет (Шварц и др., 1976). При более низкой плотности средние темпы онтогенеза повышаются, а изменчивость уменьшается (Сурова, 1985), тогда как в случае выращивания головастиков поодиночке изменчивость снова возрастает, а высокие средние темпы онтогенеза сохраняются (Сурова, 1985; табл. 3). Считается, что основным механизмом внутрипопуляционных взаимодействий являются экзометаболиты - вещества белковой природы, выделяемые головастиками в воду. Определенное значение имеют также тактильные и зрительные контакты между особями (Роус, Роус, 1964; Сурова, 1985; Шварц и др., 1976; Adolph, 1931; Gromko et al., 1973; Guyetant, 1973, 1976; Heusser, 1972; Holder, 1958; Licht, 1967, 1970 и др.). В естественных водоемах, в принципе, такая же картина (Пикулик, 1976; Северцов, Сурова, 1979а; Сурова, 1985).
Анализ наследуемости темпов роста и развития головастиков, проводимый по стандартным методикам на сибсах (Рокицкий, 1974), не дает однозначных результатов. Коэффициент наследуемости в наших исследованиях в зависимости от условий опыта колебался от h2g=0,665 до 0 (Северцов, Сурова, 1981; Сурова, 1985). В то же время изменчивость по темпам роста имеет определенные пределы, хотя внутри этих пределов темпы роста очень непостоянны. Анализ 157 индивидуальных траекторий роста головастиков показал, что ускорение наблюдается у 22% особей, а замедление- у 78%. При сохранении темпов роста неизменными отношение среднесуточных приростов (вычисленное по формуле х=l3-l2/t3:l2-l1/t1, где l - длина тела от конца морды до края анального отверстия в мм, a t - время) равно единице. В среднем это соотношение равно 0,69, т. е. темпы роста по мере онтогенеза тормозятся, хотя лимиты этого показателя колеблются от 0,06 до 3,63 (табл. 1, I, III) (Северцов, Сурова, 1981). Изменчивость среднесуточных приростов головастиков выше, чем изменчивость тех же головастиков по длине тела (табл. 1, III), что обусловлено перекрестом траекторий роста (рис. 1). Каждый головастик как бы выбирает свое место в общем распределении по темпам роста и тем самым по размерам в зависимости от своих генотипических особенностей и меняет это место в зависимости от ситуации.
| Серия | № промера | Дни от вылупления | N | lim | x | σ2 | Cυ | Стадии |
|---|---|---|---|---|---|---|---|---|
| - | 1 | 20 | 71 | 4,9-5,8 | 5,4±0,03 | 0,05±0,01 | 4,0±0,34 | 39 |
| I | 2 | 31 | 64 | 7,5-14,2 | 11,2±0,20 | 2,61±0,46 | 14,5±1,37 | 40-45 |
| I | 3 | 42 | 64 | 11,0-18,0 | 14,6±0,19 | 2,30±0,41 | 10,4±0,9 | 43-49 |
| II | 1 | 20 | 72 | 4,9-5,8 | 5,4±0,03 | 0,05±0,04 | 3,95±0,34 | 39 |
| II | 2 | 31 | 62 | 10,4-14,3 | 12,4±0,11 | 0,79±0,14 | 7,17±0,64 | 43-45 |
| II | 3 | 42 | 44 | 11,6-15,8 | 14,2±0,13 | 0,68±0,15 | 5,8-0,65 | 45-49 |
| III | - | - | 64 | -0,06-+3,63 | 0,69±0,06 | 0,26±0,04 | 74,5±3,3 | - |
Эта пластичность более четко выявляется при вычислении корреляции между начальными и конечными размерами головастиков: r=0,13 (rn=0,19, rb=0,43). Корреляция настолько мала, что невозможно говорить о фиксированных траекториях роста головастиков. Как сказано выше, опыты по содержанию головастиков в группах низкой плотности и поодиночке показывают, что они за-программированы на максимально быстрый рост при данных условиях среды. Поэтому в каждый данный момент времени нахождение любого головастика в той или иной части распределения обусловлено его физиологическим состоянием в рамках, ограниченных генотипическими возможностями. Вместе с тем в любой последующий момент времени, при любом изменении условий, например гибели части особей, изменении освещенности, питания и т. д., происходит перегруппировка генотипов внутри распределения, и в каждом размерном классе оказываются другие особи. Очевидно, эта перегруппировка может происходить только в пределах нормы реакции каждой данной особи.
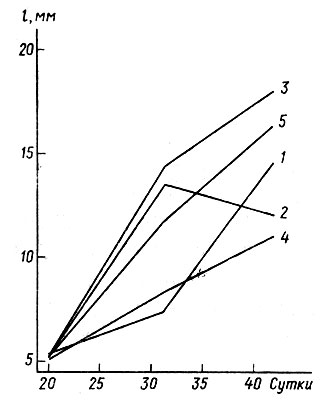
Рис. 1. 5 из 157 индивидуальных траекторий роста головастиков Ratia temporaria: 1 - наибольшее ускорение роста, 2 - наибольшее замедление роста, 3 - самый крупный головастик, 4 - самый мелкий головастик, 5 - головастик, наиболее близкий к средним для популяции темпам роста
Для того, чтобы, несмотря на недостоверность данных по наследуемости темпов роста, выяснить, влияет ли генотипический состав группы на ее фенотипическую изменчивость, был поставлен опыт с подразделением группы на части. Следует подчеркнуть, что в производных группах, представляющих собой части исходной группы (рис. 2, I-IV; табл. 2, I-IV), режим кормления, химизм среды и плотность оставались неизменными. Изменен был только один параметр - набор генотипов. Этого изменения оказалось достаточно, чтобы вызвать перераспределение головастиков внутри каждой группы. Первично единая группировка головастиков (N=379) из одной кладки икры, выращенных в лаборатории до стадии № 39 (Дабагян, Слепцова, 1975) при плотности 5 особей на 1 л воды (рис. 2, I), была разделена на 4 части (рис. 2,2). В первую вошли наиболее мелкие особи (рис. 2, 2, I), находившиеся по своему развитию в начале стадии № 39 (начало формирования почки задней конечности). Во вторую (рис. 2,2, II) вошли средние по размерам особи на той же стадии развития, что и в предыдущей группе. В третью группу (рис. 2, 2, III) были отобраны головастики тех же размеров, что и в предыдущую, но находившиеся на второй половине стадии №39 (коническая почка задней конечности). В четвертой группе находились наиболее крупные головастики, по развитию соответствовавшие второй половине стадии № 39.
Как видно из рис. 2, 2, в момент разделения распределения в дочерних группах ничем не напоминали нормальные. Однако через 10 дней все они приняли вид, свойственный любой аквариумной группировке головастиков повышенной плотности (рис.2, 3). Таким образом, в каждой дочерней группе установилось новое соотношение генотипов, обусловливающее дальнейший нормальный онтогенез каждой особи. Однако, несмотря на перегруппировку генотипов, каждая из вновь сформированных групп сохраняла свойственные ей средние темпы роста (табл. 2). Наиболее мелкие остались наиболее мелкими, наиболее крупные - наиболее крупными, а промежуточные - промежуточными. Следовательно, изменение темпов роста каждого головастика уже, чем диапазон изменчивости по этому признаку группы в делом.
| Дни от разделения групп | Группа | N | lim | x | σ2 | Сυ | Стадии |
|---|---|---|---|---|---|---|---|
| - | Исходная | 379 | 4,7-7,3 | 6,3±0,02 | 0,17±0,01 | 6,80±0,26 | 39 |
| 0 | I | 108 | 4,7-6,0 | 5,74±0,03 | 0,08±0,01 | 5,09±0,34 | 39 |
| 0 | II | 95 | 6,1-6,6 | 6,3±0,02 | 0,03±0,004 | 2,70±0,22 | 39 |
| 0 | III | 93 | 6,1-6,6 | 6,4±0,02 | 0,02±0,003 | 2,28±0,15 | 39 |
| 0 | IV | 84 | 6,7-7,3 | 6,8±0,02 | 0,02±0,003 | 2,04±0,15 | 39 |
| 10 | I | 108 | 7,2-11,4 | 8,7±0,06 | 0,35±0,05 | 6,75±0,48 | 39-44 |
| 10 | II | 95 | 7,6-10,3 | 8,9±0,03 | 0,30±0,04 | 6,12±0,44 | 39-43 |
| 0 | III | 93 | 6,7-10,4 | 8,8±0,07 | 0,45±0,07 | 7,6±0,59 | 39-43 |
| 10 | IV | 79 | 7,9-10,9 | 9,7±0,07 | 0,40±0,06 | 6,49±0,56 | 39-48 |
| 22 | I | 108 | 7,5-13,8 | 10,9+0,09 | 0,89+0,12 | 8,59+0,61 | 39-48 |
| 22 | II | 94 | 9,0-13,4 | 11,4+0,09 | 0,77+0,11 | 7,65+0,59 | 42-47 |
| 22 | III | 93 | 8,8-14,0 | 12,0±0,09 | 0,84±0,12 | 7,59±0,59 | 40-48 |
| 22 | IV | 79 | 10,3-14,0 | 12,2±0,08 | 0,47±0,08 | 5,58±0,48 | 43-48 |
| 33 | I | 100 | 8,0-12,9 | 11,3±0,09 | 0,88±0,13 | 8,28±0,57 | 40-48 |
| 33 | II | 90 | 9,1-14,4 | 11,7±0,11 | 1,02±0,15 | 8,60±0,68 | 42-48 |
| 33 | III | 80 | 9,9-13,7 | 12,3±0,10 | 0,73±0,12 | 6,89±0,56 | 43-48 |
| 33 | IV | 72 | 10,5-14,6 | 13,0±0,10 | 0,79±0,13 | 6,73±0,59 | 41-48 |
Можно, однако, предположить, что сам эффект группы моди-фицирует темпы роста сильнее, чем изменение качественного состава группы. Для проверки этого предположения был поставлен опыт по ссаживанию в группы выращенных поодиночке головастиков и рассаживанию выращенных в группе головастиков по одному (табл. 3). Как видно из таблицы, ни ссаживание, ни рассаживание на средних темпах роста не сказались. Более заметно они сказались на изменчивости по этому признаку (ср. σ2 I->II, II->I и II). Эти данные также свидетельствуют об ограниченности широты нормы реакции по темпам роста. Наконец, из 157 проанализированных индивидуальных траекторий роста был обнаружен лишь один случай столь сильного ускорения роста, что головастик, минимальный по размерам, при следующем измерении оказался третьим по длине тела. Столь же сильного торможения роста не найдено. Таким образом, темпы роста головастиков представляют собой пластичный, обладающий широкой нормой реакции признак, причем широта его нормы реакции, как и следовало ожидать, уже диапазона реакции популяции в целом. В то же время существует индивидуальная, по всей вероятности, генетически обусловленная изменчивость широты нормы реакции головастиков по темпам их роста.
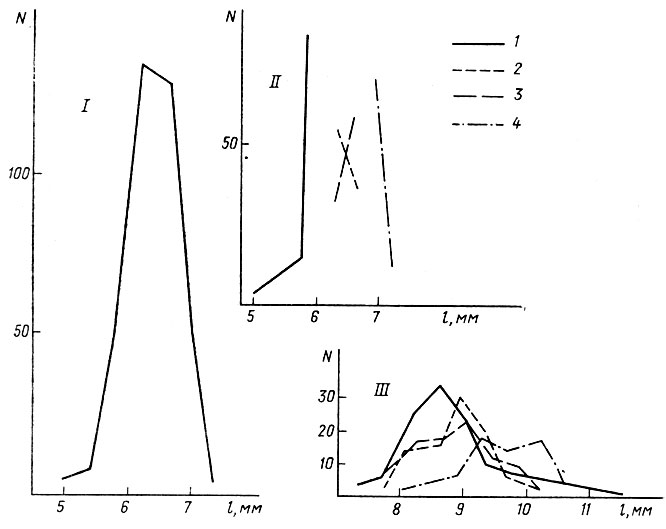
Рис. 2. Изменчивость длины тела головастиков Rana temporaria (l, мм) в зависимости от состава группы: I - вариационная кривая исходной группы, II - распределение в выделенных группах (1-4) в момент их выделения, III - распределение в тех же группах через 10 дней
Чтобы выяснить механизм действия отбора на расширение нормы реакции, представлялось целесообразным смоделировать схему Шмальгаузена: действие стабилизирующего отбора на пластичные признаки фенотипа, конкретно - на темпы роста головастиков. Для этого необходимо предварительно выяснить характер распределения головастиков по отбираемому признаку при развитии их в отсутствие элиминирующего фактора (табл. 4, 5). В аквариумы была помещена икра из одной кладки на стадии поздней гаструлы. Минимальные плотности - 45 особей на 30 л воды (табл. 4) и 50 особей на 33 л воды - соответствовали оптимальной (Шварц и др., 1976). При этой плотности средние темпы роста головастиков выше, чем при повышенной. По мере онтогенеза дисперсия по длине тела растет, причем с повышением плотности в общем быстрее, чем при оптимальной (см. табл. 1, I, II и табл. 2). Вместе с тем флуктуации изменчивости темпов роста довольно значительны. Эти флуктуации не удается связать с плотностью, поскольку все параметры содержания были выравнены, они, видимо, определяются генетическим составом группы. Форма кривых распределения головастиков по длине тела меняется в процессе онтогенеза. Как правило, она приобретает отрицательную асимметрию, выраженную тем сильнее, чем выше плотность (табл. 5). Этот эффект объясняется не гибелью быстро растущих, а увеличением числа отставших в росте головастиков, создающих "шлейф" мелких особей. Уменьшение асимметрии в последнем промере объясняется тем, что перед метаморфозом, т. е. у более крупных особей, темпы роста замедляются и кривая приобретает более симметричную форму. Показатель эксцесса (табл. 5) колеблется незакономерно, что свидетельствует об отсутствии преимущественной гибели головастиков в крайних классах распределения. Смертность составляет 12-42% от начальной численности, и не влияет на форму распределения головастиков. В естественных популяциях наблюдается аналогичная картина, также определяемая динамикой онтогенеза головастиков, но не их смертностью (Северцов, Сурова, 1979а; Сурова, 1985).
| Серия | № промера | Дни от вылупления | N | lim | x | σ2 | С0 | Стадии |
|---|---|---|---|---|---|---|---|---|
| I | 1 | 15 | 70 | 4,2-6,0 | 5,1±0,04 | 0,10±0,02 | 6,2±0,52 | 38-39 |
| I | 2 | 28 | 64 | 8,8-16,0 | 12,2±0,22 | 3,08±0,54 | 14,4±1,2 | 39-45 |
| I | 1 | 16 | 63 | 4,0-5,8 | 5,3±0,03 | 0,07±0,01 | 5,2±0,57 | 38-39 |
| I | 2 | 29 | 57 | 9,4-15,8 | 13,9±0,17 | 1,59±0,30 | 9,05±0,86 | 41-45 |
| I->II | 1 | 15 | 67 | 3,7-5,0 | 4,6±0,03 | 0,05±0,01 | 4,9±0,41 | 37-39 |
| I->II | 2 | 30 | 56 | 6,4-11,9 | 9,8±0,06 | 1,54±0,29 | 12,6±1,30 | 39-44 |
| II->I | 1 | 22 | 58 | 3,6-7,6 | 5,3±0,71 | 0,36±0,02 | 11,2±1,04 | 37-39 |
| II->I | 2 | 29 | 44 | 5,3-11,9 | 8,8±0,19 | 1,65±0,36 | 14,6±1,65 | 39-43 |
| II | 1 | 20 | 81 | 4,6-5,6 | 5,4±0,05 | 0,17±0,03 | 7,50±0,64 | 39 |
| II | 2 | 29 | 68 | 7,6-11,6 | 9,8±0,11 | 0,83±0,14 | 9,30±0,78 | 39-42 |
| II | 1 | 22 | 63 | 4,8-7,6 | 6,4±0,08 | 0,37±0,07 | 9,5±0,90 | 39 -40 |
| II | 2 | 29 | 59 | 7,6-12,0 | 9,9+0,13 | 1,03+0,19 | 10,3+0,93 | 39-44 |
Для непосредственного выяснения влияния отбора на пластичный признак в экспериментальных популяциях головастиков были созданы условия, увеличивающие их смертность. В качестве элиминирующего фактора были выбраны повышенная температура и течение воды. Группировки головастиков Rana temporaria повышенной плотности (7,5-8,5 особей на 2 л воды), происходящие из одной кладки и выравненные по стадии развития - почка задней конечности (стадии № 39) (Дабагян, Слепцова, 1975), были рассажены в аквариумы (табл. 6). Первая группировка находилась в условиях круглосуточного подогрева (+25±1°), что соответствует верхней границе температурного оптимума (Терентьев, 1950). Вторая группировка круглосуточно подвергалась воздействию кругового течения со скоростью ≈10 мм/с. Контролем служили аналогичные группировки, также выравненные по стадии развития и происходившие из той же кладки икры. Выравнивание головастиков по стадии должно было сократить изменчивость. Обе нагрузки были относительно мягкими. Отличие от естественных условий состояло только в том, что в природных водоемах подобная температура действует на скопления головастиков травяной лягушки 7-9 ч в сутки, а в опыте она поддерживалась непрерывно (Северцов, 1981б). Скорость потока воды в аквариуме не превышала скорости потока, создаваемого движением головастиков в крупном скоплении. В естественных условиях головастики двигаются мало, только при опасности, суточных миграциях и отчасти при питании. Круговое течение заставляло их непрерывно держаться против потока, расходуя на это энергию.
| Серия | Показатели | 7/V 1978 | 21/V 1978 | 1/VI 1978 |
|---|---|---|---|---|
| I. №=45 | N | 44 | 40 | 40 |
| I. №=45 | x | 6,11±0,05 | 14,7±0,12 | 16,7±0,1 |
| I. №=45 | σ2 | 0,12±0,025 | 0,58±0,12 | 0,38±0,084 |
| I. №=45 | Cυ | 5,88±0,24 | 5,22±1,19 | 3,72±0,098 |
| I. №=45 | A | 0,02±0,35 | -0,09±0,37 | -0,2±0,37 |
| I. №=45 | E | -0,1±0,69 | 0,17±0,73 | 0,13±0,73 |
| II. №=45 | N | 31 | 30 | 30 |
| II. №=45 | x | 7,2±0,07 | 16,55±0,14 | 16,48±0,17 |
| II. №=45 | σ2 | 0,15±0,038 | 0,65±0,167 | 0,85±0,219 |
| II. №=45 | Cυ | 5,5±0,21 | 4,89±0,17 | 5,6±0,22 |
| II. №=45 | A | -0,01±0,42 | -0,66±0,42 | -0,44±0,42 |
| II. №=45 | E | -0,7±0,81 | -0,36±0,82 | 0,23±0,82 |
| III. №=90 | N | 70 | 64 | 66 |
| III. №=90 | x | 6,62±0,04 | 14,4±0,09 | 15,6±0,09 |
| III. №=90 | σ2 | 0,15±0,025 | 0,51±0,09 | 0,61±0,106 |
| III. №=90 | Cυ | 6,02±0,256 | 4,96±0,17 | 5,01±0,18 |
| III. №=90 | A | -0,51±0,28 | -1,1±0,29 | -0,23±0,29 |
| III. №=90 | E | 1,07±0,56 | 1,71±0,58 | -0,21±0,58 |
| IV. №=180 | N | 120 | 110 | 104 |
| IV. №=180 | x | 6,7±0,03 | 13,1±0,07 | 14,69±0,08 |
| IV. №=180 | σ2 | 0,1±0,012 | 0,51±0,068 | 0,72±0,099 |
| IV. №=180 | Cυ | 4,78±0,16 | 5,49±0,21 | 5,0±0,18 |
| IV. №=180 | A | -1,53±0,22 | -1,43±0,23 | -0,04±0,23 |
| IV. №=180 | E | 0,05±0,43 | 2,87±0,45 | 0,34±0,47 |
Можно было ожидать, что при воздействии течения воды в первую очередь будут гибнуть наиболее мелкие, т. е. наиболее слабые, особи, и наиболее крупные, т. е. быстрее (растущие, особи, расходующие энергию на увеличение массы собственного тела.
| Серия | Показатели | 16/V 1978 | 27/V 1978 | 7/VI 1978 |
|---|---|---|---|---|
| I. №=50 | N | 50 | 40 | 40 |
| I. №=50 | x | 7,41±0,08 | 12,66±0,11 | 14,4±0,1 |
| I. №=50 | σ2 | 0,33±0,066 | 0,56±0,12 | 0,43±0,096 |
| I. №=50 | Cυ | 7,75±0,42 | 5,92±0,25 | 4,58±0,15 |
| I. №=50 | А | -0,49±0,33 | -0,17±0,37 | -0,39±0,37 |
| I. №=50 | Е | -0,46±0,66 | 0,05±0,53 | -0,17±0,73 |
| II. №=50 | N | 50 | 37 | 34 |
| II. №=50 | x | 6,99±0,06 | 13,3±0,15 | 15,3±0,1 |
| II. №=50 | σ2 | 0,19±0,038 | 0,93±0,216 | 0,4±0,097 |
| II. №=50 | Cυ | 6,23±0,27 | 7,95±0,37 | 4,14±0,12 |
| II. №=50 | А | 0,38±0,33 | 0,55±0,38 | -0,42±0,4 |
| II. №=50 | Е | -0,43±0,66 | -0,61±0,75 | -0,53±0,78 |
| III. №=50 | N | 50 | 46 | 44 |
| III. №=50 | x | 7,11±0,07 | 12,3±0,11 | 14,3±0,15 |
| III. №=50 | σ2 | 0,27±0,05 | 0,63±0,13 | 1,09±0,232 |
| III. №=50 | Cυ | 7,41±0,39 | 6,45±0,29 | 7,31±0,38 |
| III. №=50 | А | -0,02±0,33 | -0,2±0,34 | -2,0±0,35 |
| III. №=50 | Е | 0,29±0,66 | 16,69±0,68 | 4,81±0,7 |
| IV. №=50 | N | 50 | 48 | 48 |
| IV. №=50 | x | 7,25±0,07 | 12,7±0,13 | 15,0±0,12 |
| IV. №=50 | σ2 | 0,24±0,048 | 0,82±0,167 | 0,78±0,159 |
| IV. №=50 | Cυ | 6,85±0,33 | 7,15±0,36 | 5,89±0,25 |
| IV. №=50 | А | 0,17±0,33 | -2,8±0,34 | -2,02±0,34 |
| IV. №=50 | Е | -0,69±0,66 | 16,66±0,68 | 7,46±0,67 |
| V. №=50 | N | 50 | 49 | 47 |
| V. №=50 | x | 7,53±0,06 | 11,1±0,11 | 14,5±0,09 |
| V. №=50 | σ2 | 0,21±0,042 | 0,61±0,123 | 0,4±0,082 |
| V. №=50 | Cυ | 6,17±0,27 | 7,08±0,35 | 4,39±0,14 |
| V. №=50 | А | 0,04±0,33 | -0,19±0,33 | -0,28±0,34 |
| V. №=50 | Е | -0,92±0,66 | -0,24±0,6 | 0,42±0,68 |
Следовательно, лимиты распределения головастиков по размерам по сравнению с контролем должны сближаться, а эксцесс - увеличиваться. Тем самым был бы смоделирован стабилизирующий отбор.
| Серия | Показатели | 27/V 1978 | 5/VI 1978 |
|---|---|---|---|
| Подогрев t=25° | N | 149 | 88 |
| Подогрев t=25° | x | 5,65±0,02 | 7,2±0,07 |
| Подогрев t=25° | σ2 | 0,09±0,01 | 0,51±0,076 |
| Подогрев t=25° | Cυ | 5,39±0,21 | 9,86±0,69 |
| Подогрев t=25° | А | 0,01±0,19 | -0,13±0,25 |
| Подогрев t=25° | Е | -0,5±0,39 | 0,3±0,5 |
| Подогрев t=25° | lim | 5,1-6,2 | 5,2-8,9 |
| Контроль | N | 150 | 115 |
| Контроль | x | 5,46±0,03 | 7,55±0,06 |
| Контроль | σ2 | 0,18±0,02 | 0,5±0,065 |
| Контроль | Cυ | 7,82±0,43 | 9,37±0,62 |
| Контроль | А | 1,3±0,19 | -0,03±0,22 |
| Контроль | Е | 0,44±0,39 | 0,51±0,44 |
| Контроль | lim | 4,5-6,4 | 5,6-10,0 |
| Течение U=10 мм/с | N | 169 | 47 |
| Течение U=10 мм/с | x | 5,5±0,02 | 6,07±0,06 |
| Течение U=10 мм/с | σ2 | 0,11±0,012 | 0,41±0,084 |
| Течение U=10 мм/с | Cυ | 6,1±0,26 | 10,64±0,80 |
| Течение U=10 мм/с | А | 0,11±0,18 | 0,11±0,34 |
| Течение U=10 мм/с | Е | -0,5±0,37 | -1,11±0,67 |
| Течение U=10 мм/с | lim | 4,7-0,37 | 5,0-7,3 |
| Контроль | N | 156 | 111 |
| Контроль | x | 5,4±0,02 | 7,63±0,05 |
| Контроль | σ2 | 0,12±0,01 | 0,36±0,047 |
| Контроль | Cυ | 6,52±0,30 | 7,11±0,35 |
| Контроль | А | 0,86±0,19 | -0,46±0,22 |
| Контроль | Е | -0,19±0,38 | 0,16±0,44 |
| Контроль | lim | 4,4-6,1 | 5,3-9,8 |
Подогрев, стимулируя метаболизм головастиков, а отсюда темпы их онтогенеза, как и течение воды, должен стимулировать энергетические затраты. Принято считать, что само по себе развитие практически не требует дополнительных затрат энергии (Barth, Barth, 1954). По данным Л. А. Ковальчук (1978), энергетические траты на развитие у головастиков возрастают только в период метаморфоза, тогда как описываемый опыт был окончен на стадии 42, т. е. за 6 стадий до начала метаморфоза. При изучении связи энергетического обмена головастиков с их размерами Л. А. Ковальчук получила противоречивые данные: в одних случаях корреляция была положительной, в других - нулевой, в третьих - отрицательной. Можно было предполагать, что усиление энергетического обмена под действием повышенной температуры может сказаться на любом крае распределения или на обоих краях аналогично нулевой гипотезе относительно течения. Любое изменение распределения в сочетании с повышенной смертностью дало бы сведения о механизме действия отбора при ухудшении внешних условий.
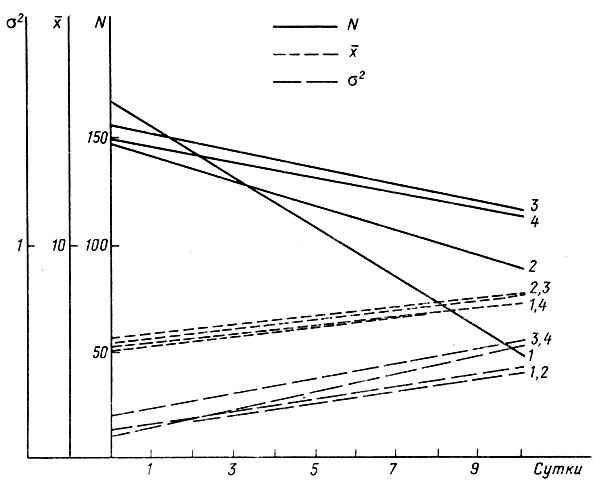
Рис. 3. Динамика численности (N), средней длины тела (х) и дисперсии по длине тела (σ2) головастиков Rana temporaria при ухудшении условий онтогенеза: 1 - круговое течение, 2 - подогрев, 3, 4 - контроль
Из рис. 3 и табл. 6 видно, что оба воздействия привели к резкому повышению смертности головастиков. В опыте с подогревом на 8-й день их осталось примерно 59%, в опыте с течением за то же время выжило лишь 28%, тогда как в контроле сохранилось 75 и 76% особей соответственно. Формы кривых распределения в обоих опытах оставались близкими к нормальной. Изменения показателя асимметрии лежат в пределах ошибки измерения. За пределы ошибки вышел показатель эксцесса, но при подогреве он увеличился, а при течении - понизился, причем то и другое в пределах колебаний этого показателя в других опытах (см. табл. 5). Изменения показателей эксцесса и асимметрии в контролях были сходны с таковыми в других опытах с повышенной плотностью. Изменчивость головастиков (табл. 6), несмотря на их высокую смертность, не понизилась по сравнению с контролем. По F-критерию различия дисперсий в опыте и контроле не достоверны. Это, как и неизменность формы кривых распределения, указывает на то, что гибель распределялась по всем классам совокупности пропорционально.
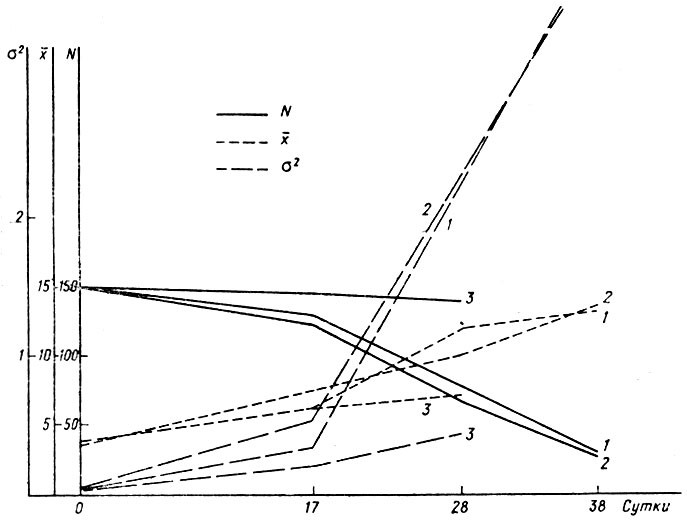
Рис. 4. Динамика численности (N), средней длины тела (х) и дисперсии по длине тела (σ2) головастиков Rana escuienta при ухудшении условий онтогенеза: 1 - круговое течение с подогревом воды, 2 - подогрев без течения, 3 - контроль
Несоответствие полученных результатов ожидаемым ставит вопрос о том, какие головастики смогли избежать гибели, т. е. о подвергавшемся отбору признаке. Ухудшение условий было общим для всей группировки, подвергавшейся данному воздействию. По своим физиологическим свойствам, в данном случае темпам роста, головастики были различны. Поэтому пропорциональная их гибель, без снижения изменчивости, может свидетельствовать лишь о том, что отбор шел не по уровню метаболизма (темпам роста), а по способности головастиков перестраивать свой метаболизм таким образом, чтобы адаптироваться к новому фактору среды и избежать элиминации. Иными словами, шел отбор особей, обладающих более широкой нормой реакции по темпам роста.
Для проверки этого вывода был поставлен близкий по методике опыт на головастиках Rana esculenta (рис. 4). Представление об отборе особей с более широкой нормой реакции подразумевает на основе вышеизложенных данных перераспределение выживших головастиков по размерным классам и тем самым подстройку их к новым условиям среды. Не способные перестроить и метаболизм, и темпы роста гибнут. Поскольку в вышеописанном опыте не было получено существенного увеличения изменчивости, этот эффект можно было только подозревать. Головастики Rana esculenta были выбраны потому, что их онтогенез протекает медленнее, чем у R. temporaria, а температурный оптимум выше. Головастики R. esculenta крупных скоплений не образуют. В контрольном аквариуме температура была +21±1°. Все это позволяло надеяться на более надежное выявление различий между опытом и контролем. Головастики, так же как и в предыдущем случае, происходили из одной кладки. Однако, поскольку величина кладок у R. esculenta меньше, выравнивания по стадиям не было проведено и контроль был один (рис. 4, табл. 7). Икра была помещена в 3 аквариума по 150 особей в каждый при плотности 8 особей на 2 л воды. Подогревали оба подопытных аквариума (t=30±1°). В одном из них, кроме того, было создано круговое течение 5 мм/с.
Результаты представлены в табл. 7. Как и в предыдущем опыте, гибель в аквариумах с усиленными физиологическими нагрузками была выше, чем в контрольном. Форма кривых несущественно отличалась от нормальной. Изменчивость же по длине тела нарастала быстрее, чем в опыте с R. temporaria. Различие между каждой из подопытных группировок R. esculenta и контрольной достоверна (Р<0,01). Таким образом, благодаря более медленному развитию R. esculenta проявился эффект, который в опыте с R. temporaria можно было только предполагать: увеличение смертности приводило к возрастанию изменчивости выживших особей и перераспределению их внутри группировки благодаря изменению траектории роста. Это перераспределение в целом не было направленным. Сохранение формы кривой свидетельствует о том, что, избегая неблагоприятных перегрузок, т. е. уравновешивая энергетические затраты на сопротивление им и на рост, головастики и ускоряли и замедляли скорость роста.
| Серия | Показатели | 20/VI 1979 | 7/VII 1979 | 18/VII 1979 | 28/VII 1979 |
|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 |
| I. Течение V=5 мм/с | N | 156 | 131 | 82 | 30 |
| Подогрев t=30 °С | x | 3,46±0,016 | 7,65±0,064 | 10,4±0,17 | 13,9±0,33 |
| Подогрев t=30 °С | σ2 | 0,04±0,005 | 0,53±0,065 | 2,27±0,35 | 3,33±0,859 |
| Подогрев t=30 °С | Cυ | 6,07±0,26 | 9,41±0,62 | 14,4±1,46 | 12,95±1,17 |
| Подогрев t=30 °С | А | -0,21±0,19 | 0,43±0,21 | 0,32±0,26 | 0,56±0,42 |
| Подогрев t=30 °С | Е | 0,17±0,34 | -0,48±0,42 | -0,95±0,52 | -0,15±0,83 |
| Подогрев t=30 °С | lim | 3,0-3,9 | 6,1-9,5 | 7,5-13,7 | 10,9-18,5 |
| II. Подогрев t=30 °С | N | 150 | 125 | 67 | 28 |
| II. Подогрев t=30 °С | x | 3,43±0,018 | 6,63±0,05 | 12,1±0,19 | 13,9±0,31 |
| II. Подогрев t=30 °С | σ2 | 0,05±0,006 | 0,33±0,042 | 2,5±0,43 | 2,66±0,71 |
| II. Подогрев t=30 °С | Cυ | 6,41±0,29 | 8,59±0,52 | 13,22±1,23 | 11,5±0,94 |
| II. Подогрев t=30 °С | А | -0,05±0,2 | 0,22±0,22 | -0,12±0,29 | -0,32±0,44 |
| II. Подогрев t=30 °С | Е | -0,16Η0,39 | -0,47±0,43 | -0,66±0,57 | -1,07±0,85 |
| II. Подогрев t=30 °С | lim | 2,9-4,0 | 5,5-8,2 | 8,3-15,4 | 10,9-17,4 |
| III. Контроль | N | 150 | 146 | 133 | - |
| III. Контроль | x | 3,67±0,02 | 6,5±0,04 | 7,2±0,06 | - |
| III. Контроль | σ2 | 0,06±0,007 | 0,2±0,023 | 0,44±0,054 | - |
| III. Контроль | Cυ | 6,81±0,32 | 6,92±0,34 | 9,16±0,59 | - |
| III. Контроль | А | -0,19±0,2 | -0,11±0,2 | 0,24±0,21 | - |
| III. Контроль | Е | -0,18±0,39 | -0,47±0,40 | -0,53±0,41 | - |
| III. Контроль | lim | 3,1-4,2 | 5,4-7,4 | 5,8-9,3 | - |
Едва ли следует думать, что при столкновении организма с воздействием внешней среды, столь сильным, что оно может вызвать гибель данного организма, не будут включаться все возможные для данного организма реакции, способные хоть в какой-то степени снизить вероятность гибели. Трудно предполагать, что изменение темпов роста - единственный или даже последний из механизмов, обеспечивающих выживание головастиков при ухудшении их физиологического состояния. Но механизм этот действует и действует эффективно. В этом отношении можно утверждать, что выжившие головастики прошли отбор по широте нормы реакции при данной силе физиологических нагрузок. В череде поколений такой отбор приведет к расширению нормы реакции в целом. Предлагаемый механизм отбора на расширение нормы реакции получен в лабораторных опытах. Поэтому необходимо было показать, что он действует и в естественных популяциях. Для этого был выбран подход, аналогичный вышеизложенному: попытка смоделировать стабилизирующий отбор, но уже на головастиках, взятых из естественных водоемов.
Одним из наиболее известных и общепризнанных примеров действия стабилизирующего отбора в природе является работа Г. Бумпуса (Bumpus, 1899), изучавшего гибель воробьев, окоченевших во время метели в Нью-Йорке. Результат этого исследования, неоднократно перепроверенный (Jonston et al., 1972; O'Donald, 1973 и др.), сводится к тому, что наиболее устойчивыми к резкому понижению температуры оказались особи с морфологическими параметрами, близкими к модальному (среднему) значению, тогда как погибли особи, уклоняющиеся и в плюс-, и в минус-направлениях. Аналогичные результаты, подтверждающие эффективность действия стабилизирующего отбора, идущего по стабильным признакам фенотипа, получены на ряде видов животных (Curtzinger, 1976; Fretwell, 1977; Mason, 1969; Scheiring, 1977).
Мы попытались промоделировать стабилизирующий отбор на головастиках Rana temporaria, используя резкие кратковременные воздействия, интенсивность которых подбирали таким образом, чтобы получить смертность, близкую к Ld=50. Как и в работе Бумпуса, учитываемым признаком служили линейные размеры - длина тела от конца морды до анального отверстия, хотя в описываемых опытах, как и в случае с воробьями, элиминирующие воздействия влияют не на морфологию, а на физиологию. Отличие состояло только в том, что размеры тела и его частей у взрослых воробьев - признаки стабильные для данного животного, тогда как размер тела головастиков - признак, меняющийся во времени, т. е. характеризующий не только физиологическое состояние, но и изменения самого этого признака. Физиологическое состояние головастиков в водоемах (как и в аквариумных группировках) определяется тремя противоречивыми тенденциями: затратами на поддержание жизни; генетической программой, заставляющей головастиков расти и развиваться с максимальной Для данных условий среды скоростью, и ингибирующим рост влиянием группы. Насколько противоречивы две последние тенденции, видно из сравнения данных по росту головастиков, воспитываемых поодиночке (в 0,7 л воды каждый) и в группе, в которой на каждую особь приходилось такое же количество воды (Сурова, 1982). За первые 30 дней после вылупления среднесуточный рост "одиночек" составлял 0,433 мм, а у головастиков в группе "оптимальной" плотности - лишь 0,327 мм.
Для моделирования стабилизирующего отбора в качестве элиминирующих факторов были взяты: эфирный наркоз, воздействие нейтральным красным и температурный шок (Северцов, 1984). Устойчивость к эфирному наркозу может характеризовать лишь общее физиологическое состояние организма и только через это состояние может быть связана с темпами роста. Нейтральный красный подавляет деление клеток (Ромейс, 1953), в малых концентрациях - тормозит рост головастиков (Travis, 1981). Температурный шок для пойкилотермных головастиков наиболее точно имитировал резкое воздействие холодом на гомойотермных воробьев. Во всех трех случаях (табл. 8) выборки выживших и погибших не отличаются, значения χ2 при P=0,05 меньше статистических. Это значит, что гибнут особи любых размерных классов.
| Серия | Показатели | Эфирный наркоз | Нейтральный красный | Тепловой шок |
|---|---|---|---|---|
| Выжившие | N | 107 | 130 | 125 |
| Выжившие | x | 10,5±0,03 | 9,0±0,03 | 10,6±0,02 |
| Выжившие | lim | 6,1-14,9 | 5,5-12,6 | 6,3-14,9 |
| Выжившие | σ2 | 3,11 | 2,87 | 3,07 |
| Выжившие | стадии | 39-47 | 39-47 | 39-47 |
| Погибшие | N | 96 | 97 | 102 |
| Погибшие | x | 10,5±0,03 | 8,5±0,01 | 11,0±0,04 |
| Погибшие | lim | 5,7±15,4 | 5,2±11,9 | 6,3±15,8 |
| Погибшие | σ2 | 3,05 | 1,75 | 4,13 |
| Погибшие | стадии | 39-47 | 39-45 | 39-47 |
| - | χ2вычисл | 6,7 | 5,9 | 10,57 |
| - | К | 6 | 5 | 6 |
| - | χ2статист | 12,59 | 11,07 | 12,50 |
Можно было бы предполагать, что рост головастиков не связан с выбранными воздействиями, тем более что они очень кратковременны: эфирный наркоз в концентрации 30 мл на 1 л воды в течение 10 мин, 0,002%-ный нейтральный красный в течение 5 мин и подъем температуры от 17 до 35° за 5 мин при тепловом шоке. При таких условиях возможна только физиологическая реакция. Но темпы роста, как показано выше, тесно связаны с физиологическим состоянием, и, следовательно, резистентность выживших головастиков определялась оптимальным для каждой данной особи соотношением затрат на темпы роста и остального энергетического обмена. Иными словами, погибли те головастики, чьи темпы роста в течение предшествующей жизни были таковы, что привели к ухудшению их физиологического состояния. Поскольку распределения выживших и погибших головастиков не отличаются друг от друга, данный опыт, подтверждая опыты с дополнительными физиологическими нагрузками, свидетельствует о том, что в природе отбор на расширение нормы реакции связан с гибелью особей всех классов распределения по отбираемому признаку, не способных в силу узости своей нормы реакции адаптироваться к элиминирующему воздействию. Использованные воз-действия, выявляя головастиков, резистентных к ним, тем самым выявляют головастиков, толерантных к предшествующей экологической обстановке.
Полученные результаты кардинально отличаются от результатов работы Г. Бумпуса и аналогичной работы С. Фретвелла (Fretwell, 1977). Это можно объяснить тем, что темпы роста в отличие от дефинитивных мерных признаков могут меняться во времени. Если у воробьев адаптация к холодовому шоку достигалась только за счет физиологических реакций, на которые темпы роста влияли в прошлом, что и обусловило пониженную устойчивость уклоняющихся особей, то у головастиков устойчивость к элиминирующим воздействиям определялась различной степенью оптимизации взаимодействия обоих параметров. Поэтому в отличие от воробьев судьба головастиков зависит не от близости их размеров к среднему значению, а от сбалансированности темпов роста и физиологического состояния независимо от размеров данной особи, т. е. от широты нормы реакции каждого из двух параметров.
Из вышеизложенного следует, что механизм отбора на расширение нормы реакции существенно отличается от предлагавшегося И. И. Шмальгаузеном стабилизирующего отбора, влияющего на крайние классы распределения (см. выше). На самом деле при ухудшении внешних условий (которые всегда сопровождают их флуктуации) в любой части распределения, а не только на его краях, выживают особи, способные уклоняться от данного воздействия. Увеличение индивидуальной изменчивости при ухудшении условий обитания отмечено многими авторами (Агаев, 1978; Ежиков, 1933; Поляков 1975; Филипченко, 1978). Это подтверждает описанный механизм отбора на расширение нормы реакции. Более прямым подтверждением вышесказанного могут служить результаты опытов по дизруптивному отбору количественного признака - массы тела у Drosophila melanogastev (Higuet, 1986). Подобный отбор приводит лишь к резкому увеличению изменчивости мух по этому признаку. Так как в данном случае дизруптивный отбор велся путем сохранения крайних вариантов, т. е. против существующей адаптивной нормы, можно утверждать, что его структура - селективное преимущество уклонений - была близка к структуре отбора на расширение нормы реакции.
Поскольку гибель, причиной которой служит недостаточная приспособленность по пластичным признакам фенотипа, вызывается в значительной степени случайными констелляциями неблагоприятных обстоятельств внешней среды, ее часто обозначают как неизбирательную элиминацию. Подобное представление возникает из-за множественности действия внешних факторов (Шварц, 1969; Шилов, 1977). Однако гибель от таких констелляций факторов идет в природе постоянно, а так как в популяции гибнут не все особи (подобную смерть нельзя считать неизбирательной), отбор на расширение нормы реакции идет постоянно.
Фактор (или комплекс факторов), вызывающий наиболее интенсивную элиминацию, будет приводить к наиболее интенсивному расширению нормы реакции, т. е., если речь идет об онтогенетических реакциях, к выработке адаптивных модификаций. Именно поэтому адаптивные модификации представляют собой реакции на комплекс внешних воздействий, или, что то же самое, в эксперименте (и в природе) данная модификация может быть вызвана разными воздействиями. Поэтому одно и то же воздействие среды может вызвать комплексную реакцию организма, и разложить комплексность воздействий и комплексность реакций организма на элементарные парные взаимодействия с однозначным соответствием, как правило, трудно или невозможно.
Например, на темпы роста головастиков влияют температурный режим, состав пищи и обеспеченность ею, зрительные и тактильные взаимодействия особей данной популяции, химизм воды и ряд других факторов. В природе все они действуют в совокупности, хотя в каждый данный момент времени наиболее сильное действие оказывает один или немногие факторы. Необходимостью адаптации к комплексу внешних факторов объясняется и то, что одну и ту же адаптивную модификацию можно в эксперименте вызвать разными воздействиями. Так, известная модификация - окраска крыльев у бабочек Arachinia (Vanessa) levana-prorsa L., отражающая способность к перенесению куколками зимнего сезона, - может быть вызвана охлаждением куколок, любым другим воздействием, вызывающим диапаузу, а также световыми воздействиями на куколку (Яхонтов, 1969). Уменьшение длины крыльев у дрозофилы при повышении температуры до +28° происходит и за счет уменьшения средних размеров клеток, и за счет уменьшения их числа (Алпатов, 1935). Согласно Бонье (цит. по Филипченко, 1978), горная модификация одуванчиков Toraxacum densleonis отличается от равнинной не только размерами (горная форма меньше), но и изменениями формы листьев, относительным удлинением корня, относительным укорочением цветоносов и рядом других признаков. Аналогично у позвоночных физиологические реакции, например суточная активность, регулируются эндогенно (циркадный ритм), фотопериодом, температурными изменениями и в ряде случаев звуковой сигнализацией.
Существование геномных или морфогенетических корреляций между признаками (Шмальгаузен, 1982) не подразумевает того, что и селективная ценность таких признаков коррелирована, напротив, одни из плейотропных проявлений мутаций могут иметь положительную селективную ценность, другие - отрицательную. Множественность элиминирующих факторов приводит к усилению гибели по любому, недостаточно пластичному из коррелированных между собой признаков и к расширению нормы реакции всего их ансамбля. Это означает приобретение каждым из признаков селективной ценности, сходной с селективной ценностью остальных, коррелированных с ним признаков. По мере расширения нормы реакции всех признаков увеличивается селективная ценность каждого из них, тогда как реакции организма становятся более целостными.
Другим свойством адаптивных модификаций да и вообще пластичных признаков фенотипа, получающим объяснение с вышеизложенных позиций, является наличие скрытого резерва адаптивного реагирования, проявляющегося только в ответ на действие экстремальных для данного вида факторов среды. Например, акклиматизация в высокогорье (3000-4000 м) равнинных пород овец, крупного рогатого скота, лошадей сопровождается сначала повышением интенсивности дыхания, затем, после возвращения частоты дыхания к норме, увеличением количества гемоглобина, в частности числа эритроцитов, и далее, при снижении гемоглобина, - интенсификацией гликолиза (Слоним, 1962). Параллельно снижается гормональная активность щитовидной железы, что свидетельствует об общем понижении уровня метаболизма (Курбанова, 1979). Едва ли акклиматизируемые в горах равнинные животные когда-либо сталкивались со столь существенным снижением парциального давления кислорода, но адаптироваться к подобным, эволюционно новым факторам среды они способны. Ряд аналогичных примеров был использован Л. С. Бергом (1977, с. 246-254) для обоснования непосредственного формообразующего действия ландшафта, т. е. для доказательства недарвиновского механизма эволюции.
Наличие скрытого резерва адаптивного реагирования трудно-объяснимо с позиций концепции И. И. Шмальгаузена (см. выше). С позиций предлагаемой концепции оно находит объяснение. Отбор на расширение нормы реакции приводит в череде поколений к формированию непрерывного спектра онтогенетических, физиологических или поведенческих реакций, позволяющих особям уклоняться от элиминации. При наличии воздействия включается тот или иной онтогенетический (или физиологический) механизм, что и приводит к формированию соответствующей реакции, в том числе адаптивной модификации. Иными словами, внешнее воздействие, вызывающее соответствующую реакцию, с самого начала имеет до некоторой степени сигнальное значение: направление изменения среды определяет направление реакции. Однако, поскольку для выживания важна именно пластичность реагирования, пределы выражения реакции генетически не запрограммированы. Напротив, отбор на расширение нормы реакции все время повышает пластичность признака. Поэтому более сильное или более продолжительное, чем обычно, действие средового фактора автоматически приводит к более сильному проявлению реакции, в конечном итоге к выявлению скрытого резерва адаптивного реагирования. Так, равнинные породы скота вынуждены приспосабливаться к недостатку кислорода не потому, что его концентрация в атмосфере заметно колеблется, а потому, что в зависимости от энергозатрат колеблется потребность в нем организма. Оказавшись в высокогорье в условиях недостатка кислорода, они используют уже существующие компенсаторные механизмы, которые в новых условиях работают непрерывно и с большей нагрузкой, вызывая дальнейшие изменения метаболизма, а затем отбор на фиксацию данного механизма как основного.
Скрытый резерв адаптивного реагирования обусловлен кроме непосредственного расширения нормы реакции под действием отбора сложной системой обеспечения надежности поддержания гомеостаза организма. В первую очередь, это множественное обеспечение биологически важных функций (Маслов, 1980), позволяющее организму адекватно реагировать даже на очень глубокие изменения и внешней среды, и состояния других систем организма (патологию). Во многом это достигается благодаря наличию систем обеспечения структурной надежности (Федоров, 1988). Так, для обеспечения нормальной работы главного глазодвигательного нерва достаточно 4•103 нейтронов, на самом деле их имеется 2,5•104. Для обеспечения работы синоатриального узла сердца надо примерно 10 клеток, тогда как в этом узле их приблизительно 10 тыс. Число аналогичных примеров может быть увеличено. Отчасти подобная "избыточность" связана с условиями функционирования самих компонентов системы - необходимостью их физиологической репарации - и временного прекращения выполнения главной функции (Федоров, 1988), например периодический переход в фазу покоя пролиферирующих кроветворных клеток и пристеночное стояние моноцитов и гранулоцитов.
Однако в значительной степени существование большого количества компонентов функциональных систем, не объяснимое исходя из условий функционирования подобных систем, связано с необходимостью обеспечения гомеостаза при повышении функциональной нагрузки. Данные, собранные В. И. Федоровым, показывают, что патологические состояния организма, вызванные как инфекционными заболеваниями, так и мутационными нарушениями, создают гораздо более сильные функциональные нагрузки на соответствующие системы организма, чем изменения абиотических или биотических условий внешней среды. Это означает, что отбор на расширение нормы реакции, вызванный патологическими состояниями, должен быть наиболее интенсивным. Очень интересной в этом отношении представляется вероятность существования отбора на расширение нормы реакции, вызванного необходимостью компенсации мутационно обусловленных патологических состояний. Такой отбор может служить механизмом превращения неблагоприятных мутаций в селективно-нейтральные. Тогда накопление какой-то их части оказывается процессом, подчиняющимся не стохастическим закономерностям, постулируемым теорией нейтральности, а селективным.
Реакции пластичных признаков фенотипа на изменения среды будут адаптивными до тех пор, пока возможно нормальное функционирование механизма, обеспечивающего ее проявление, а само проявление будет повышать вероятность выживания и (или) размножения. Поскольку нормальное функционирование поддерживается всем гомеостазом организма, а реакция с самого начала адаптивна, морфозы и патологические состояния при неэкстремальных воздействиях среды - явление, по-видимому, достаточно редкое. К подобным не адаптивным морфофизиологическим реакциям можно отнести радиационные и хемоморфозы рыб и амфибий, возникающие при соответствующем загрязнении окружающей среды, талидомидовый морфоз развития рук у человека и т. п. Нарушение целесообразности адаптивности реакций без существенных нарушений гомеореза и (или) гомеостаза организма также может встречаться в природе. Например, у бабочки крапивницы (Vanessa urticae L.) при действии на нее сверхвысокими (42-46°) или сверхнизкими (-20-0°) температурами возникает одна и та же абберация - морфа cohnusoides. Эта морфа иногда встречается в природе на южной границе ареала вида (Fischer, 1930, цит. по Светлову, 1978).
Отсутствие четких пределов адаптивных реакций организма и постоянство действия отбора на расширение нормы реакции ставит вопрос о пределах, до которых она может расширяться, и о факторах, обусловливающих эти пределы. Очевидно, что по мере расширения нормы реакции отбор в этом направлении будет становиться все менее эффективным из-за снижения наследуемости отбираемого признака. Кроме того, как уже сказано, чем дальше заходит реакция организма на внешнее воздействие, тем вероятнее возникновение неадаптивных проявлений ее или патологических состояний, нарушающих гомеостаз (или гомеорез) организма, включая морфозы, стресс и т. п. Подобные слишком сильно укло" няющиеся организмы подвергаются более интенсивной элиминации, что ведет к стабилизирующему отбору по широте нормы реакции, т. е. к фиксированию лимитов изменений при модификации. Тот же механизм действует и при вторичном сужении нормы реакции, происходящем в тех случаях, когда вид сталкивается с более узким диапазоном колебаний внешних условий, и часть возможных проявлений его нормы реакции становится неадаптивной.
Это показано К. Бервеном и Д. Джилл (Berven, Gill, 1983) при изучении наследуемости темпов роста у головастиков Ranu silvatica в популяциях с равнин Мэриленда, горных популяций Виргинии и тундровых -Северной Канады. В горных и северных популяциях наследуемость темпов роста выше, чем в равнинных: равнина - 0,08±0,004, горы - 0,58±0,17, тундра - 0,27±0,05, что объясняется краткостью теплого сезона. Повышение коэффициента наследуемости означает возможность повышения эффективности отбора и, следовательно, темпов эволюции пластичных признаков фенотипа. В данном случае следует четко различать темпы эволюции и темпы адаптации. Темпы эволюции пластичных признаков фенотипа могут быть очень низкими из-за низкого (если не нулевого) коэффициента наследуемости подобных признаков и необходимости генокопирования для продолжения отбора по подобным признакам. В то же время темпы адаптации особей и популяций при помощи подобных признаков, как показал И. И. Шмальгаузен (1968а), могут быть очень высокими, вплоть до приспособления к новым условиям за одно поколение. Таким образом, границы адаптивного реагирования обычно шире, чем это наблюдается в пределах колебаний внешних условий, но все же ограничены стабилизирующим отбором неадаптивных и нарушающих целостность организма реакций.
Как уже говорилось, отбор на расширение нормы реакции приводит к выработке реакций, в том числе и модификаций, с непрерывным спектром проявления. Дальнейшая эволюция подобных адаптаций сводится к трем основным вариантам: 1) смещению нормы реакции при изменении внешних условий; 2) выработке дискретных модификаций в пределах нормы реакции; 3) дальнейшему совершенствованию реакции на воздействия среды.
Механизм смещения нормы реакции при изменении внешних условий, по-видимому, соответствует представлениям И. И. Шмальгаузена (1968а). Проявления нормы реакции, не совпадающие с изменениями среды, элиминируются, что должно приводить к ее одностороннему сужению, но одновременно по адаптивным проявлениям действует отбор на ее расширение. Форма кривой распределения по отбираемому признаку зависит от широты нормы реакции и степени изменения среды за единицу времени. Если норма реакции большей части особей в популяции перекрывает диапазон изменений внешнего фактора, распределение смещается без существенного изменения своей формы, если не перекрывает, то, как это показал Г. Д. Поляков (1975), кривая принимает асимметричную форму: большая часть особей меняет свой фенотип в соответствии с изменением среды, не способные же полностью адаптироваться к новым условиям особи составляют шлейф изменчивости, направленный в сторону, противоположную направлению отбора. По мере селекционной адаптации к новым условиям этот шлейф должен исчезать, что будет соответствовать "стабилизации нормы" по Шмальгаузену (см. выше).
Примерами смещения нормы реакции может служить известная работа Н. И. Калабухова и А. М. Родионова (1936) об адаптации Apodemus silvaticus к существованию в горах. При завозе в Кавказский заповедник мышей, пойманных в равнинной части Ставропольского края, у них повышалось количество гемоглобина, не достигая, однако, уровня, свойственного горным популяциям этого вида. В случае перемещения "горных" мышей на равнину количество гемоглобина у них снижалось, но оставалось выше равнинной нормы. Таков же смысл вышеупомянутого примера с фиксацией модификации листьев у Апаетопае Pulsatilla.
Возникновение дискретных адаптивных модификаций И. И. Шмальгаузен связывал с действием стабилизирующего отбора в разнородно меняющейся среде. Действительно, такие модификации, как три формы листьев стрелолиста (Sagittaria sagittifolia), адаптивны к трем дискретным средам: лентовидные - подводные, округлые - плавающие и стреловидные - воздушные. Этому предположению противоречит, однако, большое число дискретных адаптивных модификаций, приспособленных к непрерывному градиенту факторов среды. У того же стрелолиста форма листьев определяется освещенностью почек в предыдущем сезоне, освещенность же - фактор не дискретный. Переключение креодов двух дискретных модификаций Osmerus eperlanus L. обусловлено не дискретностью их сред обитания, а окупаемостью (или не окупаемостью) энергетических затрат на рост. Если энергетические затраты растущей рыбы на добывание крупной добычи оправдываются, рост продолжается и получается корюшка, имеющая длину примерно 17 см. Если же затраты не оправдываются, рост прекращается, происходит половое созревание и возникает снеток - планктоноядная рыбка длиной 6-10 см (Криксунов, Шату. новский, 1979). Количество пищи так же фактор не дискретный.
Приведенные примеры свидетельствуют, что возникновение дискретных модификаций из непрерывной паратипической изменчивости обусловлено не дискретностью сред, в которых проявляются модификации, а совершенствованием механизмов реагирования на внешние воздействия. Формирование дискретных путей развития обеспечивает более надежное соответствие проявлений нормы реакции данным условиям среды. К предлагаемой схеме креодизации модификационной изменчивости близок, по представлениям Г. М. Длусского (1981), путь возникновения полиморфизма рабочей касты у муравьев. Возникновение, точнее, выделение дискретных модификаций из непрерывного их спектра представляет собой не что иное, как креодизацию развития (в смысле Уоддингтона), не связанную в данном случае с его эквифинальностью. Переход от одного креода к другому - переключение креодов, как видно из приведенных примеров (число которых можно увеличить), обусловлено сигнальным действием средовых факторов, а сами креоды разграничены порогами реагирования развивающихся систем (см. ниже). Возможно, аналогичные принципы, основанные на внешнем по отношению к данному формообразовательному аппарату воздействии, ответственны за переключение креодов при эквифинальном развитии, характерном для нормального морфогенеза многоклеточных.
Не менее (если не более) важным направлением эволюции пластичных признаков фенотипа является эволюция темпоральных (временных) характеристик реакций организма. Любое изменение поведения, физиологических реакций, а тем более онтогенеза, требует времени на его реализацию. Оно представляет собой именно реакцию - ответ организма на внешнее воздействие и реализуется с запозданием по отношению к началу воздействия. Особи, реакции которых не успевают за изменениями среды, должны подвергаться более интенсивной элиминации. Поэтому для адаптации на основе пластичных признаков фенотипа крайне важны синхронизация или упреждение реакциями организма изменений внешних условий. Роль синхронизации реакций организма и изменений реакций среды подчеркнул И. А. Шилов (1985). То, что подобное запаздывание адаптации действительно существует, можно видеть на примере тех реакций, для которых сроки реализации не имеют выраженного селективного значения: либо благодаря мультиобеспеченности соответствующих функций (Маслов, 1980), либо благодаря нейтральности самого признака. Например, согласно Ю. Б. Бызовой (1977), у ряда беспозвоночных полная адаптация газообмена к повышению температуры происходит за 7-14 дней, хотя наиболее сильные его изменения наблюдаются в первые дни. Так как интенсификация обмена обеспечена многими факторами, такими как гликолиз, использование кислородного резерва тканей и т.д., запаздывание реакции, видимо, не влечет элиминации. Увеличение размеров тела у парамеций при их культивировании в условиях пониженной температуры обусловлено замедлением клеточного деления (Алпатов, 1935). К повышению элиминации это не ведет, так как более низкие темпы прироста численности компенсируются снижением энергетических затрат при возрастании объема.
В ряде случаев само запаздывание реакции относительно вызывающего его воздействия адаптивно. Так, любое изменение темпов роста и развития головастиков при изменении их плотности (см. выше) адаптивно до тех пор, пока сохраняется данная плотность населения. Аналогично изменение окраски у одиночной и стадной фаз саранчи, обусловленное также эффектом группы, обеспечивает существование в соответствующих условиях плотности и количества корма. Однако во многих случаях запаздывание реакции по отношению к внешнему воздействию, даже если оно не влечет за собой гибели, может сказываться на относительной при-способленности или участии в размножении. Например, в плохо прогреваемых водоемах замедление темпов онтогенеза головастиков Rana temporaria на фоне эффекта группы приводит к тому, что отставшие в развитии особи не успевают метаморфизировать до наступления холодов (Северцов, Сурова, 1979а). Но при большей пластичности темпов развития тот же элиминирующий фактор может приводить, напротив, к замедлению онтогенеза и зимовке головастиков, как это наблюдается у Pelabates fuscus. Иными словами, при эволюции признаков с широкой нормой реакции в определении направленности эволюции на первый план выступает не изменчивость, подвергающаяся отбору в данных условиях, а, как при филогенезе (см. раздел II), организация признака. В данном случае - организация онтогенеза.
Представление о неблагоприятных последствиях запаздывания реакций организма, по-видимому, не требует специальных доказательств. С этих позиций можно рассматривать даже успех (или неуспех) хищника при нападении на жертву. Если жертва поймана, то ее поведенческие реакции были недостаточно быстрыми, если же жертва избежала нападения, то недостаточно быстрыми были реакции хищника.
Смягчению неблагоприятных последствий запаздывания реакций служат многие механизмы, включая множественное обеспечение биологически важных функций - мультиобеспеченность (Маслов, 1980), многоуровневость разных по скорости адаптивных реакций (Северцов, 1922) и сама скорость этих реакций. Однако большая часть этих механизмов не способна синхронизировать изменения организма и среды, а тем более обеспечить упреждение внешних воздействий. Поэтому важным способом адаптации, снижающим или устраняющим неблагоприятные воздействия запаздывания реакций, является выработка порогов реагирования. Возможно, превращение плавно развивающейся реакции в пороговую представляет собой неизбежный путь дальнейшего совершенствования любых адаптивных реакций. На это указывает огромное число примеров пороговых реакций, начиная с реакций тканей и клеток на гормональные воздействия и кончая различными, например температурными, адаптациями целостного организма. Так, у сумеречных и ночных животных активность начинается при достижении некоего, видоспецифичного порога освещенности. Например, у сипухи Tuto alba и ушастой совы Asio otus максимальная активность наблюдается при освещенности 0,4 лк, а у домового сыча Athene noctua-150 лк. Малые подковоносы вылетают на охоту при освещенности 20-150 лк (Шилов, 1985). Поскольку освещенность зависит от многих факторов (времени года, облачности, экспозиции, закрытости биотопа и т.д.), такой механизм обеспечивает тонкую подстройку периода активности к благоприятному времени суток.
Адаптивность пороговых реакций по сравнению с плавно раз-вивающимися обусловлена тем, что подпороговые значения внешних факторов не вызывают изменений - организм к ним резистентен. По достижении порогового значения воздействия реакция соответствующей системы реализуется полностью, перекрывая и по скорости, и по степени изменения весь диапазон действия внешнего фактора или приводя фенотип в соответствие этому фактору в пределах генотипических возможностей организма. Достаточно ярким примером, иллюстрирующим адаптивное значение порогов реагирования, служит пространственная и временная суммация нервных импульсов в синапсах нервных ганглиев насекомых. От-носительно поведения примером пороговых реакций может служить частота контактов между животными. Достижение определенной частоты подобных контактов служит сигналом к миграции, снижающей плотность населения (Шилов, 1977).
Видимо, основным путем становления и развития пороговых реакций является изменение чувствительности к средовому воздействию в результате отбора. Чем более "эволюционно освоено" - термин Шмальгаузена - подобное воздействие, т. е. чем дальше зашла по отношению к нему адаптация, тем резче выражена адаптивная реакция. Так, согласно Н. Г. Лопатиной (1971), выработка пищевых условных рефлексов у пчел по отношению к запаху обычно посещаемых ими растений требует минимального числа (одно-двух) подкреплений, после чего кормушку посещают 95-100% пчел. Выработка аналогичной реакции на редко посещаемые растения происходит дольше и требует серии из 6-7 подкреплений. Еще труднее (примерно за 10 подкреплений) вырабатывается условный рефлекс на запах искусственного пищевого объекта (бутиловый спирт). Предпосылкой для выработки пороговых реакций служит то, что факторы среды, вызывающие ту или иную модификацию, с самого начала имеют сигнальное значение, на что указывают многочисленные данные по акклимации. Акклимация, не меняя состояния организма, облегчает его дальнейшую адаптацию к изменениям среды (Freda, Dunson, 1986). Подчас эти адаптации имеют очень сложный, опосредованный характер. Так, по данным В. В. Васильевой (1986), температурный отбор спермиев у лягушек приводил к повышению теплоустойчивости эпителия жабр головастиков, вылупившихся из икры, оплодотворенной теплоустойчивыми спермиями.
Иногда пороговые реакции оказываются недостаточно эффективными. В этих случаях вырабатываются сигнальные реакции, часто сочетающиеся с пороговыми. Сигнальные реакции получают решающее значение, если по условиям механизма своего осуществления или гомеостаза организма ответ на внешнее воздействие развивается относительно медленно, или, что то же самое, изменение внешних условий происходит относительно быстро, а реакция на него длительна. Тогда норма реагирования признака, запускающего адаптивную реакцию, сужается еще сильнее, и внешний фактор получает сигнальное значение. В основе этого направления эволюции пластичных признаков фенотипа лежат главным образом два вышеупомянутых свойства пластичных признаков фенотипа: реакция организма на внешние воздействия комплексна, а потому, в сущности, делима на серию более или менее чувствительных к внешнему фактору частных реакций; внешний фактор с самого начала адаптации к нему имеет сигнальное значение.
При отборе на сигнальное значение внешних воздействий норма реакции признака, наиболее чувствительного к этому воздействию и запускающего весь комплекс частных реакций, сужается, и внешний фактор становится сигналом (для поведенческих реакций - релизером). Происходит это, видимо, при отборе на снижение пороговой чувствительности или, может быть, непосредственно при отборе на повышенную чувствительность, минуя этап пороговых реакций. Многие физиологические и поведенческие реакции, такие как суточные и годовые физиологические циклы, а также более специфические реакции, например линька и сезонные миграции, чрезвычайно сложны и включают биохимические, эндокринные, гистологические, поведенческие и другие процессы, обычно регулируемые гипоталамо-гипофизарной системой и высшими центрами мозга (Шилов, 1985). Одна из наиболее детально исследованных подобных реакций - миграционное состояние птиц (Дольник, 1975). Рассмотрение процессов эволюционного формирования таких реакций затруднено их сложностью и недостатком сравнительных данных. В общей форме оно требует введения представления о косвенном отборе (соотборе), т. е. об отборе на коадаптацию компонентов подобных систем. При таком подходе представление о направленности эволюции каждого компонента системы может быть сведено к вышеизложенному представлению об отборе по единичному признаку, обладающему (или не обладающему) той или иной степенью пластичности.
Однако, по-видимому, решающее значение в формировании и дальнейших преобразованиях подобных систем имеют онтогенетические, в частности эргонтические корреляции (Шмальгаузен, 1938). Раз возникнув, связь между компонентами системы обусловливает затем взаимосвязанные их изменения. Так как все эти компоненты обладают некой широтой нормы реакции, эволюционный механизм преобразований сложных систем оказывается достаточно простым и сводится к ситуации отбора на расширение или сужение нормы реакции компонентов системы, тогда как в целом она сохраняется, лишь перестраиваясь в соответствии с требованиями экологии данного вида. Так, при сохранении сложнейших взаимодействий циркадного ритма и фотопериодических реакций в качестве общей основы сезонной регуляции физиологического состояния организма у суслика Spermophilus lateralis, преимущественно зерноядного, годовой цикл контролируется в основном эндогенно, а у S. beldingi, питающегося суккулентами, - средовыми стимулами (Шилов, 1985).
Сигнальность внешних воздействий позволяет синхронизировать начало изменения окружающей среды и реакции организма. Например, известно, что теневые и световые модификации листьев древесных растений обусловлены степенью освещенности почек в предшествующем сезоне вегетации. Поэтому величина листовых пластинок не зависит от изменений их освещенности по мере роста листьев в данном сезоне, а предвосхищает окончательные значения освещенности. Иными словами, сигнальность позволяет упреждать начало изменений внешней среды. Однако сами факторы среды, к которым организм приспосабливается посредством пластичных признаков фенотипа, как правило, не стабильны ни по времени действия, ни по силе. Иначе не нужны были бы адаптации к ним через расширение нормы реакции. Вот почему в ходе дальнейшей эволюции сигнальное значение приобретают не те воздействия внешней среды, к которым адаптивна данная реакция, а сопутствующие ей более стабильные внешние факторы, например, фотопериод - фактор астрономический, а потому наиболее стабильный, или более или менее стабильные проявления других параметров. Так, В. Р. Дольник (1975) показал, что эволюция физиологических механизмов миграции у птиц шла по пути замены тех экологических факторов, к которым она адаптивна (неблагоприятный сезон, бескормица и т.п.), фотопериодическими реакциями, подготавливающими миграционное физиологическое состояние и стимулирующими саму миграцию. Изменения фотопериода стимулируют и диапаузу у насекомых, и многие другие реакции (Рощупкин, Потапенко, 1977). Развитие сигнальных реакций является, по-видимому, наиболее эффективным способом упреждения организмом неблагоприятных воздействий среды. Особенно эффективен такой способ реагирования для наиболее быстрых поведенческих реакций. Недаром именно поведенческие реакции позволяют предвосхищать изменения среды на основе опережающего отражения (Анохин, 1968) или экстраполяционного рефлекса (Крушинский, 1977).
Таким образом, эволюция пластичных признаков фенотипа,, и в частности адаптивных модификаций, происходит под действием трех основных компонентов естественного отбора: отбора на расширение нормы реакции, который по своей структуре напоминает не стабилизирующий, а балансирующий, для одного поколения- центробежный отбор (Симпсон, 1948) и приводит к формированию пластичных признаков фенотипа; стабилизирующего отбора, ограничивающего или сужающего норму реакции, и стабилизирующего отбора на пороговые и (или) сигнальные значения внешних факторов, в результате которого происходит стабилизация одного из компонентов комплексной реакции, позволяющая синхронизировать изменения организма и внешней среды и в ряде случаев упреждать их.
В целом рассмотрение эволюции пластичных признаков фенотипа вносит лишь одно изменение в представление о направленности эволюции подобных признаков под действием отбора. При изменении внешней среды направленность преобразования организации проявляется гораздо сильнее, чем при движущем отборе, идущем по генотипическим вариациям фенотипов. В случае адаптации посредством пластичных признаков сначала происходит изменение, а затем уже генотипическая фиксация этого изменения. При других формах отбора - стабилизирующем и балансирующем (во всех их проявлениях, включая отбор на расширение нормы реакции) - можно говорить о направленности эволюции не столько в смысле направленности преобразований организации, сколько в смысле адаптации, которая может выражаться в сохранении уже существущих приспособлений, а также в изменении или поддержании уже имеющегося в популяции разнообразия фенотипов и их реакций на внешние воздействия. В этом отношении эволюция признаков всегда направленна. Преобразование организации - не более чем частный случай направленности эволюции как процесса адаптации.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'