1.3. Популяция под действием системы векторов отбора
В естественных условиях любая популяция находится под действием множества факторов среды (каждый из которых может вызвать гибель или устранение от размножения части составляющих ее особей), т. е. под действием серии одновременно влияющих на нее векторов отбора. Поэтому особи каждой популяции в природе вынуждены адаптироваться одновременно в нескольких направлениях. В приложении к экологии методами математического моделирования эту ситуацию наиболее обстоятельно рассмотрел Ф. Н. Семевский (Семевский, 1979; Семевский, Семенов, 1982). Ф. Н. Семевский указал, что В. Метью (Mattew, 1926), а затем К. Кермак (Kermack, 1954) рассмотрели множественную адаптацию на конкретном материале. Дж. Холдэн (Haldane, 1956) сформулировал принцип, согласно которому в множестве состояний фенотипов адаптация в каком-либо одном направлении идет за счет ослабления адаптации в других направлениях. Под множеством состояний фенотипов (множеством Метью - Кермака, по терминологии Семевского) понимается не популяция как таковая, а множество возможных изменений (реакций) фенотипов, составляющих популяцию. Критерием оптимальности служит число половозрелых потомков, оставляемых данной особью, т. е. ее репродуктивный успех. Дж. Холдэн рассмотрел эту ситуацию для стационарной среды. Н. Семевский обобщил критерий Холдэна относительно нестационарных сред, усреднив по времени репродуктивный потенциал особи. Поэтому, видимо, справедливо принцип адаптации популяций к множеству факторов среды называть принципом Холдэна-Семевского.
Как уже сказано, суть принципа сводится к тому, что при фиксированном ресурсе и в отсутствие прогрессивной эволюции адаптация в одном каком-либо направлении идет в ущерб адаптации в других направлениях. Ресурс при этом, естественно, перераспределяется, т. е. доля его, идущая на адаптацию к рассматриваемому фактору, увеличивается, а к остальным - уменьшается. Полная адаптация к данному фактору невозможна именно из-за ограниченности ресурса. Поскольку рассматривается экологическая ситуация, в качестве механизмов адаптации обсуждаются главным образом фенотипические реакции: изменения поведения, изменения хода онтогенеза и т. п. Организмы сначала используют наиболее эффективные из доступных им способов адаптации, если этих механизмов недостаточно, включаются все другие (см. 1.2). Чем сильнее действие фактора среды, тем недостаточнее адаптивный ответ на него. В результате меняются факторы смертности: при адаптации к данному фактору на первый план выступают другие факторы среды, влияющие на другие признаки организма. Происходит смена факторов, лимитирующих численность популяции.
Принцип Холдэна - Семевского, по сути дела, представляет собой экологическое объяснение правила минимума. Экологическая его интерпретация должна дополнять и углублять Либиховскую - физиологическую интерпретацию, так как последняя оказывается трудно применимой к реальным ситуациям, наблюдаемым в природе. Действительно, почти любой фактор среды, ограничивающий расселение видов, говоря шире, возможность существования их на данной территории, сам по себе преодолим. Таковы климатические факторы, сила действия которых обычно нарастает по градиенту от зоны оптимума к зоне минимума. Таковы же и обеспеченность пищей, убежищами и т. п. На возможность эффективной адаптации к подобным факторам в их критических для вида концентрациях указывает, в частности, наличие скрытого резерва адаптивного реагирования. Поэтому способность или неспособность особей данного вида существовать при соответствующей силе действия лимитирующего фактора будет определяться не столько физиологическими возможностями адаптации именно к этому фактору, сколько взаимодействием его через перераспределение ограниченного ресурса со всеми другими действующими на популяцию факторами и продолжительностью их действия.
Проверяемыми следствиями действия принципа Холдэна - Семевского, по мнению Ф. Н.Семевского и С. М. Семенова (1982), может служить уменьшение (возможно, замедленное увеличение. - А. С.) смертности по мере усиления давления фактора среды и относительное увеличение смертности, вызываемой этим фактором, при его ослаблении. Авторы подчеркивают, что, не располагая прямыми доказательствами, считают вышеизложенное гипотезой. В обоснование ее кроме математических выкладок они приводят косвенные данные: множественность и взаимное замещение факторов смертности, таблицы выживания ряда видов, пока-зывающие ограниченность смертности от каждого средового фактора.
Прямые подтверждения принципа Холдэна - Семевского будут приведены ниже. Но независимо от них надо подчеркнуть, что логически он безупречен. Для того чтобы он выполнялся, достаточно трех условий: 1) многокомпонентности факторов среды, действующих на организм; 2) наличия коррелированности частей организма; 3) ограниченности хотя бы одного ресурса среды. Два первых условия самоочевидны и не нуждаются в доказательстве. Третье - практически общепризнано, причем в гораздо более жесткой форме. Представление об ограниченности ресурсов среды и возникающей на этой основе конкуренции лежит в основе практически всех экологических и эволюционных обобщений. Из признания ограниченности хотя бы одного ресурса следует признание конкуренции за этот ресурс, а в силу целостности организма и лимитированность реагирования по всем остальным параметрам: среды. Из лимитированности реакций следует в свою очередь недостаточность адаптации к каждому из компонентов среды. Принцип Холдэна - Семевского признается мною как реально отражающий механизм экологической адаптации популяций к комплексу совокупно действующих условий среды.
Однако для целей данной работы необходимо рассмотреть эволюционные следствия принципа Холдэна - Семевского. В эволюционном аспекте этот принцип превращается в представление о несовершенстве адаптации, или, что то же самое, в представление об относительности органической целесообразности. Как известно, это представление лежит в основе дарвинизма (Шмальгаузен, 1969; Северцов, 1981а). Поэтому вслед за экологической оптимизацией будет следовать эволюционная, происходящая посредством естественного отбора особей, наиболее адекватно реагирующих на весь комплекс средовых факторов и в первую очередь на давление наиболее сильного из них. Возникновение новых или совершенствование старых адаптаций будет оптимизировать общее соотношение организма со средой, так как снижение смертности от наиболее сильно действующего фактора (а в этом и состоит экологический смысл любого приспособления) приведет к перераспределению лимитированного ресурса на другие направления приспособления.
Из принципа Холдэна - Семевского следует, что многокомпонентность адаптации приводит к отбору по каждому из направлений взаимодействия организма со средой, если это взаимодействие достаточно сильно, чтобы вызвать элиминацию. Эффективность отбора в каждом данном направлении будет зависеть от полноты адаптации ко всем остальным факторам среды или от силы давления каждого из них (см. ниже).
С точки зрения направленности эволюции принцип Холдэна - Семевского подразумевает, что для каждого конкретного направления отбора справедлива вышеизложенная схема движущего отбора по признаку, будь этот признак стабильным или пластичным. Для организма в целом необходимо рассматривать взаимодействие всех векторов отбора. В силу их неизбежной разнонаправленности, обусловленной многокомпонентностью среды и взаимозаменяемостью факторов смертности, суммарное направление эволюции можно представить как равнодействующую всех векторов отбора. Эта равнодействующая ни по величине, ни по направлению не будет совпадать ни с одним из частных векторов. Учет коррелированности частей организма осложняет, но не меняет принципиально эту схему. Другими словами, направление эволюции популяции задается наличной изменчивостью (в данном случае всей ее совокупностью) и экологической ситуацией, к которой популяция должна адаптироваться.
Вышеизложенный подход, полностью вытекающий из принципа Холдэна - Семевского, не позволяет судить о реальном взаимодействии системы векторов отбора в естественных популяциях. В рамках этого подхода взаимодействие в рассматриваемой системе лишь постулируется на основе косвенных данных. Кроме вопроса о направленности эволюции на основе представления о множественности действующих в популяции векторов отбора можно обсуждать и вопрос о темпах микроэволюции. Действительно, со времен Ч. Дарвина обсуждается вопрос, почему новый вид, возникнув, сохраняется затем в стационарном (или квазистационарном) состоянии неопределенно долгое время?
Общеизвестны подсчеты Дж. Симпсона (Simpson, 1952, 1953), согласно которым средняя продолжительность существования видов у позвоночных составляет примерно 2,75 (0,5-5) млн лет (Gill, Cobban 1973; Stanley, 1985 и др.). Столь же общеизвестны многочисленные случаи быстрого формообразования и видообразования. Например, возникновение в течение 26 лет в Калифорнии расы яблонной плодожорки (Laspeyresia pomonella), поражающей грецкий орех (Essing, 1931); возникновение за 10 тыс. лет 18 видов рыб рода Barbus от одного исходного вида в оз. Лаоно (Майр, 1968); возникновение всего за 1000 лет 5 эндемичных видов бабочек на Гавайях (Zimmerman, 1960) и др. Несоответствие реальной неравномерности темпов эволюции и восходящей к Дарвину идеи ее медленности и постепенности породило концепцию прерывистого равновесия (Gould, Eldredge, 1977).
Не вдаваясь в детали дискуссии между градуалистами и пунк-туалистами, все же приходится признать, что обе эти концепции отражают крайние точки зрения и поэтому страдают односторонностью. Нельзя отрицать лежащего в основе градуалистических представлений медленного и плавного видообразования (микроаккумулятивное видообразование по Завадскому, 1968), доказанного многими исследователями. Наиболее яркими примерами, подтверждающими его, являются общеизвестные ряды Paludina (Viviparus) neymayri и Planorbis multiformis или океанического палеопланктона. Также нельзя отрицать и вышеупомянутые случаи быстрого видообразования, подтверждающие представления пунктуалистов (Kaplan, 1985; Stanley, 1985). Таким образом, приходится признать очень широкий диапазон скоростей эволюции. На одном его краю находится неоформогенное видообразование по К. М. Завадскому (1968), при котором изоляция возникает за одно поколение, а затем на основе обособившейся (изолированной) "новой видовой формы" образуется новый вид. Примером подобного видообразования может служить возникновение алло- и аутополиплоидов у растений, способных к вегетативному размножению. На другом краю находятся формы с очень медленным и постепенным видообразованием вплоть до персистентных форм, у которых, несмотря на индивидуальную изменчивость, фенотипический облик не меняется в течение геологических эпох. По-видимому, по ходу филогенеза темпы эволюции данной популяции также могут меняться в очень широких пределах, что и соответствует представлению о прерывистом равновесии. Следовательно, в объяснении нуждается не только высокая скорость эволюции, по и сохранение видом стационарного состояния.
Наиболее общепринятым объяснением стабильности видов является теория стабилизирующего отбора (Шмальгаузен, 1968а). Действительно, если признак сохраняет свое приспособительное значение, то до тех пор, пока условия среды для него неизменны, он будет подвергаться стабилизирующему отбору. В результате популяция по этому признаку остается неизменной. Если предположить, что, пока вид существует в свойственной ему экологической нише, большинство его признаков сохраняет приспособительное значение, можно считать, что стабилизирующий отбор ответствен за стационарное состояние видов в природе.
Подобное объяснение наталкивается, однако, на существенное затруднение. Адаптация к тому или иному внешнему воздействию подразумевает снижение смертности, причиняемой этим воздействием. Например, адаптация насекомых к инсектицидам снижает эффективность применения последних. Поэтому если большая часть признаков вида, сохраняя свое адаптивное значение, подвергается только стабилизирующему отбору, то элиминироваться, как и считал И. И. Шмальгаузен (1969), должны лишь особи,, уклоняющиеся за пределы популяционной нормы.
Согласно "классической гипотезе" организации генотипа, дикий тип гомозиготен по большинству нормальных доминантных аллелей, а выщепление рецессивных гомозигот, которые и элеминируются, - событие редкое. Согласно "балансовой гипотезе" дикий тип адаптивен в силу своей высокой гетерозиготности, и неадаптивные сочетания генов также редки. Иными словами, и с точки зрения приспособленности, и с точки зрения генотипической изменчивости элиминация при стабилизирующем отборе не должна быть интенсивной, а численность популяций, находящихся в стационарном состоянии, должна быть стабильной и высокой.
В природе подобные ситуации не наблюдаются. Напротив, смертность за одно поколение у любого вида чрезвычайно высока (ср. Семевский, Семенов, 1982). Именно смертностью за поколение со времен Ч. Дарвина принято характеризовать напряженность борьбы за существование. Так же интерпретировал ее и И. И. Шмальгаузен (1969). Например, у травяной лягушки до половой зрелости доживает не более 0,1% особей от начальной численности генерации. Если, следуя теории стабилизирующего отбора, представить себе, что популяции этого вида существуют в благоприятных для них условиях среды, придется признать, что лишь 0,1% особей нормальны по своему генотипу, а остальные 99,9% попадают в зону безусловной элиминации. Из этого в свою очередь следует, что любое изменение условий среды приведет данный вид к вымиранию, чего на самом деле не наблюдается. Вид существует по меньшей мере с плейстоцена, откуда известен скелет лягушки, не отличающийся от современных (Терентьев, 1950). Род Rana известен в Европе с верхнего эоцена (Rage, 1984). Таким образом, возникает вопрос, как сочетаются жесткий естественный отбор, на который указывает высокая смертность, и: эволюционная стабильность природных популяций. Существование подобного отбора ставит вопрос о направленности эволюции популяций, точнее, о причинах отсутствия этой направленности в период стасиса. Материалом для решения этих, вопросов послужили данные по экологии развития Rana temporaria L. Все наблюдения проводились в окрестностях г. Москвы, на Звенигородской биологической станции МГУ.
Среди бурых лягушек травяная является наиболее плодовитым и рано размножающимся видом. Из табл. 9 видно, что R. temporaria нерестится примерно на 10 дней раньше, чем ближайший к ней и симпатричный с ней в средней полосе России вид - Rana arvalis. Нерест R. temporaria начинается, как только лед отходит от берегов постоянных водоемов и образуются временные водоемы в заполняемых талыми водами ямах, колеях и т. п. Часто можно видеть пришедших на нерест травяных лягушек и их икру подо льдом, намерзшим за время ночных заморозков. Таким образом, нереститься раньше, чем это делает R. temporaria, невозможно. Все раннее развитие этого вида приходится на период, когда почва еще только оттаивает и вода в нерестовых водоемах холодная. Вместе с тем, как видно из табл. 9, продолжительность развития от икрометания до метаморфоза у травяной и остромордой лягушек примерно одинаковая, несмотря на то что развитие остромордой происходит в более мягких температурных условиях.
| Год | Rana temporaria | Rana arvalis | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| t начала икрометания, °С | икрометание | метаморфоз | время развития в воде, сут | t начала икрометания, °С | икрометание | метаморфоз | время развития в воде, сут | |||||
| даты | сутки | даты | сутки | даты | сутки | даты | сутки | |||||
| 1981 | 4,9 | 28-30/IV | 2 | 13/VI-13/VII | 30 | 76 | 14,3 | 2- 10/V | 8 | 14/VI-18/VII | 34 | 77 |
| 1982 | 5,5 | 18-29/IV | 11 | 26/VI-29/VII | 33 | 102 | 14 | 28/IV-8/V | 10 | 23/VI-14/VIII | 52 | 108 |
| 1983 | 5,7 | 9-14/IV | 5 | 16/VI-20/VII | 34 | 102 | 7 | II-26/V | 15 | 15/VI-10/VIII | 56 | 91 |
Плодовитость R. temporaria по сравнению с R. arvalis выше. Согласно П. В. Терентьеву (1950), травяная лягушка мечет 1115-4005 икринок, остромордая - 1000-2000. По нашим данным, в окрестностях Звенигородской биостанции модальная величина кладки травяной лягушки - 1700 икринок (lim 700-6700 шт.), а остромордой - 1100 (lim 280-2100 шт.). Более высокая плодовитость травяной лягушки объясняется тем, что она мечет икру во временные водоемы и на мелководьях постоянных водоемов, уровень в которых падает по окончании таяния снега. В результате и икра, и головастики часто гибнут от высыхания. Как видно из табл. 10, во временных водоемах гибнет почти вся икра. Гибель 15% кладок, отложенных в постоянные водоемы, также объясняется главным образом их обсыханием.
| Показатели | Временные водоемы | Постоянные водоемы |
|---|---|---|
| Количество водоемов | 23 | 8 |
| Количество и процент кладок, отложенных в данный тип водоема | 510 (15,7%) | 2741 (84,3%) |
| Количество и процент кладок, погибших в данном типе водоема | 448 (87,8%) | 431 (15%) |
Таким образом, существует довольно мощный вектор отбора на увеличение плодовитости травяной лягушки. И. И. Шмальгаузен первым показал, что неизбирательная гибель ведет к отбору на плодовитость. Объяснение высокой плодовитости R. temporaria неустойчивостью гидрологического режима тех водоемов, в которых нерестится этот вид, дали С. С. Шварц с соавторами (1976). С высокой плодовитостью связаны и размеры тела. Средняя длина тела половозрелых R. temporaria в окрестностях Звенигородской биостанции - 79,67±0,57 мм, a R. arvalis - 54,67±0,79 мм, т. е. на 2,5 см меньше. Корреляция размеров тела и плодовитости показана для ряда видов Anura, в том числе и для Rana temporaria (Barker 1981; Kadel, 1977; Koskela, Pasanen, 1975). Корреляция размеров икринок и плодовитости отсутствует (Сурова, устн. сообщ.).
| Температура, °С | 9/IV | 11/IV | 12/IV | 13/IV |
|---|---|---|---|---|
| Воздуха | 9,2 | 8,5 | 7,2 | 2,6 |
| Воды | ||||
| поверхность | 7,9 | 5,9 | 6,4 | 3,7 |
| дно | - | - | 6,2 | 3,8 |
| Скопления кладок | ||||
| поверхность | 9,1 | 8,3 | 7,4 | 4,0 |
| дно | - | - | 6,8 | 4,2 |
| х отдельных кладок N=10 | 8,4 | 6,3 | 6,2 | 3,9 |
Возникает вопрос, почему отбор на плодовитость и увеличение размеров, несмотря на продолжающуюся гибель от обсыхания икры и личинок, не приводит к дальнейшему увеличению среднего количества икринок в "ладке и коррелятивно размеров самих лягушек. Иными словами, почему вид R. temperaria остается стабильным. Дело в том, что существует ряд векторов отбора, противодействующих отбору на плодовитость. Как уже сказано, травяные лягушки нерестятся на оттаивающих раньше других и наиболее прогреваемых участках водоемов. Это приводит к тому, что масса кладок оказывается сгруппированной на небольшой акватории. Кладки сливаются друг с другом, образуя сплошные, часто многослойные "подушки". Это имеет адаптивное значение, так как скопления икры накапливают тепло (табл. 11). Аккумуляция тепла ускоряет эмбриональное развитие, компенсируя дефицит температуры. Следует добавить, что более высокая температура в скоплениях икры привлекает лягушек, которые мечут свою икру в те же скопления (Сурова, Северцов, 1984). У R. arvalis столь плотных скоплений икры не наблюдается.
| Даты учета и стадии развития | Верхний слой | Нижний слой | ||
|---|---|---|---|---|
| живые | погибшие | живые | погибшие | |
| 29/IV | N=1910 | N=2220 | ||
| Гаструла - хвостовая почка | 97,9% | 2,1% | 73,9% | 26,1% |
| 1/V | N=1300 | N=2260 | ||
| Нейрула - хвостовая почка | 92,3% | 7,7% | 55,8% | 44,2% |
| 3/V | N=1250 | N=699 | ||
| Хвостовая почка - вылупление | 90,4% | 9,6% | 40% | 60% |
| Всего к началу вылупления | 94,2% | 5,8% | 61,4% | 38,6% |
Однако чем больше и плотнее скопления икры, тем выше эмбриональная смертность. Даже в отдельно лежащих кладках R. temporaria всегда гибнет 10-20 центральных икринок. В скоплениях икры этого вида эмбриональная смертность составляет 20-30% (табл. 12, 13), а иногда она достигает 70% и более. Объясняется это, по-видимому, гипоксией (Сурова, Северцов, 1984). Из табл. 12 видно, что смертность нарастает по мере развития и в нижних слоях скопления икры она выше, чем в верхних. Отбор на снижение чувствительности к гипоксии, видимо, шел. На это указывает само наличие "подушек" икры, отсутствующих у R. arvalis. Но продолжение адаптации в этом направлении невозможно из-за неэффективности отбора на устойчивость к недостатку кислорода. Дело в том, что при увеличении выживания эмбрионов будет возрастать смертность на личиночных стадиях развития.
| Дата учета | Возраст в сутках и стадии развития | Численность икры и личинок | Процент гибели от исходной численности | Процент гибели за время между учетами |
|---|---|---|---|---|
| 21/IV | 0 | 630 000 | 0 | 0 |
| 26/IV | 5 начало вылупления | 447 000 | 29 | 29 |
| 2/V | 11 вылупление окончено | 288 000 | 54,2 | 36 |
| 7/V | 16 начало расплывания | 202 200 | 68 | 30 |
| 10/V | 19 массовое расплывание | 180 000 | 71,4 | 11 |
После вылупления головастики некоторое время остаются на поверхности скопления икры, питаясь ее оболочками. В это время они образуют чрезвычайно плотные агрегации, смертность в которых составляет 30 и более процентов от числа вылупившихся особей (табл. 13). Объясняется это тем, что с началом личиночного развития начинает действовать эффект группы (см. выше): чем ниже эмбриональная смертность, тем выше плотность головастиков на поверхности "подушки" икры и тем выше постэмбриональная смертность.
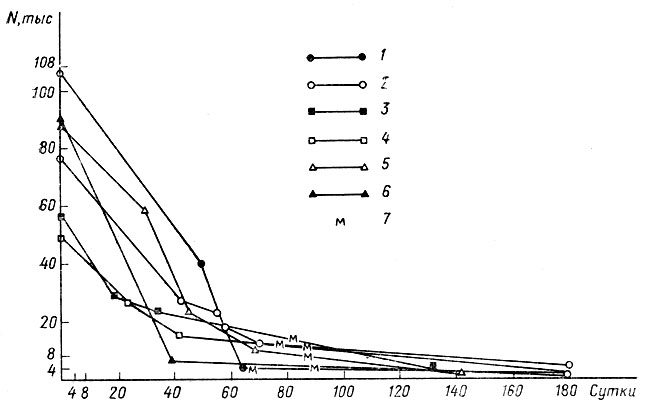
Рис. 5. Динамика численности 6 генераций головастиков Rana temporaria в одном водоеме: 1-6 - годы (соответственно с 1975 по 1980), 7 - начало метаморфоза
По достижении стадии почки задней конечности (ст. 39, по Дабагян, Слепцовой, 1975) головастики покидают места нереста и расплываются по водоему. Однако смертность их на этих (средних и поздних) стадиях личиночного развития (ст. 40-50) сохраняет зависимость от плотности. На рис. 5 показана динамика численности головастиков в одном из водоемов Звенигородской биостанции с низкой начальной численностью генерации. Даже в этих условиях зависимость смертности от плотности достаточно наглядна. Еще более наглядна она в водоемах с высокой начальной численностью генерации (рис. 6).
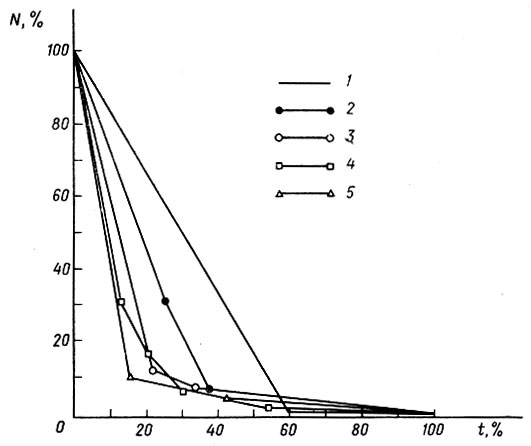
Рис. 6. Динамика численности 5 генераций головастиков Rana temporaria в водоеме с высокой начальной плотностью этих животных, в процентах от начальной численности генерации (Ni) за процент времени до начала метаморфоза (t): 1-5 - годы (соответственно с 1975 по 1979)
Предполагаемая И. Петранкой (Petranka, 1985) равномерность убыли головастиков во времени не согласуется с нашими данными. Некоторый намек на равномерность убыли можно получить при условии исчисления гибели в процентах от численности при предыдущем ее учете. При подобных расчетах смертность составляет 30-50% за каждый период онтогенеза, но падает к метаморфозу и во время этого периода. Реальная убыль численности зависит от плотности.
Хищники не оказывают влияния на изменение численности го-ловастиков. Как видно из рис. 7, подъем численности хищных беспозвоночных (главным образом это личинки плавунцов) не сказывается на кривой численности головастиков (Северцов, Сурова, 19796). Пищевая конкуренция, насколько это удалось выяснить, у головастиков отсутствует. Хотя они, по-видимому, и испытывают некоторый дефицит белковой пищи, но восполняют его поеданием трупов своих собратьев. Таким образом, на всем протяжении личиночного развития смертность определяется преимущественно внутрипопуляционными взаимодействиями. Из табл. 14 видно, что в эксперименте даже слабое повышение плотности (согласно С. С. Шварцу и др., 1976, плотность 1 особь на 0,6 л воды оптимальна) повышает и смертность, и изменчивость головастиков.
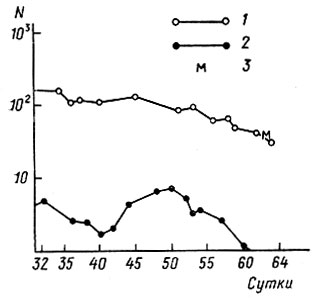
Рис. 7. Динамика плотности (N) головастиков Rana temporaria и уничтожающих их хищников в одном водоеме. По оси абсцисс - сутки с момента вылупления головастиков: 1 - головастики, 2 - хищники, 3 - начало метаморфоза
Вместе с тем после расплывания с мест нереста плотность головастиков при расчете на водоем в целом не превышает, а обычно ниже оптимальной, однако и дифференциация популяций по темпам онтогенеза, и смертность продолжаются. Это связано с тем, что головастики R. temporaria на стадиях 40-50 постоянно собираются в скопления на наиболее прогреваемых, освещенных солнцем мелководьях, у берегов водоемов. На рис. 8 показаны ход суточных температур и концентрация головастиков на учетных площадках южной и северной экспозиции. Скопления головастиков аккумулируют тепло, что необходимо им для поддержания темпов развития. В подобные скопления собираются до 70% особей, обитающих в водоеме, и иногда подобные скопления сохраняются за счет накопленного тепла даже после захода солнца. Головастики "упакованы" в этих скоплениях максимально плотно, они касаются друг друга, благодаря чему продолжает действовать эффект группы, обусловливающий как дифференциацию генерации по темпам онтогенеза, так и смертность. Поэтому чем выше численность головастиков в водоеме, тем крупнее скопления и сильнее сказывается эффект группы независимо от плотности населения, рассчитанной на водоем в целом.
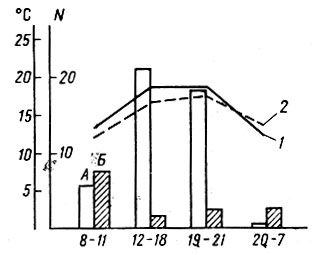
Рис. 8. Зависимость плотности головастиков от температуры среды в течение суток на площадке южной (А) и северной (Б) экспозиции: 1 - температура воздуха, 2 - температура воды
Таким образом, чем ниже эмбриональная смертность, тем выше численность головастиков в водоеме, крупнее их скопления и выше смертность на стадиях личиночного развития. Следовательно, отбор на устойчивость к гипоксии эмбриональных стадий не эффективен: повышая выживаемость эмбрионов, он автоматически приведет к повышению смертности головастиков, т. е. к уничтожению результатов увеличения приспособленности эмбриональных стадий развития. Вместе с тем эмбриональная и личиночная смертность совместно делают неэффективным и отбор на плодовитость. Чем больше отложено икры, тем выше совокупная гибель на эмбриональных и постэмбриональных стадиях и менее эффективно дальнейшее увеличение плодовитости. В принципе, существуют два пути увеличения плодовитости: увеличение числа икринок при неизменности их размеров, что связано с ростом размеров лягушек, и увеличение числа икринок за счет уменьшения каждой из них. Этот путь не связан с размерами лягушек. Возможно, этот путь был отчасти использован в эволюции R. temporaria: распределение икры по ее диаметру у травяной лягушки в отличие от остромордой бимодально (Сурова, устн. соообщ.). Лимиты диаметров икринок у обоих видов практически совпадают: R. temporaria - 2,95-4,15 мм, R. arvalis - 2,95-4,05 мм. Но у R. arvalis модальное значение признака совпадает со средним - 3,5 мм, а у R. temporaria две моды - 3,0 мм и 3,5 мм. Однако у обоих видов корреляция между диаметром и числом икринок в кладке отсутствует. Это заставляет искать иные объяснения бимодальности распределения у травяной лягушки, например наличие возрастной изменчивости или связь с условиями зимовки. В любом случае отбор на уменьшение размеров икринок не может быть эффективен, так как связан с уменьшением количества желтка, а следовательно, с ускорением эмбрионального развития, чего на самом деле не наблюдается.
| Вид и число особей | Возраст, сут | 1 особь на 0,6 л | 1 особь на 0,4 л | ||||||
|---|---|---|---|---|---|---|---|---|---|
| N | x | σ2 | Cυ | N | x | σ2 | Cυ | ||
| Rana | 13 | 85 | 6,60 | 0,15 | 5,83 | 87 | 6,16 | 0,18 | 6,96 |
| temporaria | 29 | 85 | 11,07 | 0,60 | 6,47 | 84 | 10,19 | 0,88 | 9,21 |
| N=100 | 39 | 78 | 13,23 | 0,75 | 6,53 | 78 | 17,35 | 1,66 | 11,40 |
| - | сеголетки | 71 | 10,97 | 0,55 | 6,76 | 23 | 10,80 | 1,30 | 10,56 |
| Rana | 14 | 73 | 6,63 | 0,16 | 6,09 | 78 | 6,21 | 0,20 | 7,18 |
| arvalis | 28 | 73 | 12,27 | 0,81 | 7,34 | 78 | 10,50 | 3,56 | 7,96 |
| N=100 | 40 | 67 | 14,28 | 0,94 | 6,78 | 70 | 12,44 | 3,25 | 14,50 |
| - | сеголетки | 49 | 10,77 | 0,87 | 8,68 | 7 | 10,84 | 1,70 | 12,01 |
Можно, однако, предположить, что описанная ситуация должна вызвать отбор на снижение чувствительности к эффекту группы. Второй способ избежать плотностно-зависимой гибели - отбор на изменение поведения, т. е. отбор на исчезновение скоплений, - невероятен, так как в этом случае головастики не будут получать достаточного количества необходимого для их развития тепла. Отбор на снижение чувствительности к эффекту группы у головастиков Rana temporaria, по-видимому, шел. Из табл. 14 следует, что у R. arvalis дифференциация по темпам онтогенеза, их общее замедление и смертность при повышении плотности проявляются сильнее, чем у R. temporaria. Однако, как и в случае отбора на устойчивость к гипоксии, отбор на устойчивость к эффекту группы не привел к исчезновению этого эффекта.
Чтобы выяснить, почему не действен отбор против эффекта группы, необходимо проанализировать другую особенность размножения R. temporaria: сроки икрометания и связанные с ними сроки метаморфоза. Из табл. 8 видно, что не только нерест, но и метаморфоз у травяной лягушки начинается раньше, чем у остромордой. В этом, собственно, и состоит адаптивный смысл скоплений и икры, и головастиков, компенсирующих недостаток тепла, обусловленный ранним икрометанием. Для Rana temporaria ранний метаморфоз имеет и приспособительное значение: чем раньше сеголетки травяной лягушки покидают водоем, чем крупнее они, тем жизнеспособнее (Ляпков, 1986). Поскольку темпы роста сеголеток имеют тенденцию сохраняться (Ляпков, 1986; Смирина, 1980), более мелкие и вышедшие позднее сеголетки обычно не успевают до зимовки догнать по своим размерам более крупных. Эффект группы приводит к тому, что первыми выходят на сушу наиболее крупные сеголетки, затем по мере метаморфоза генерации средние размеры особей уменьшаются, а изменчивость по этому признаку возрастает. К концу выхода сеголеток их средний размер снова несколько увеличивается (Ляпков, Смирина, 1983).
Таким образом, сеголетки, первыми покидающие водоем, по-видимому, имеют селективное преимущество и, следовательно, должен существовать вектор отбора, действующий против эффекта группы. Такой отбор способствует формированию более раннего и более синхронного метаморфоза генерации. Кроме того, как показано на рис. 6, пик численности хищников, питающихся головастиками, приходится на вторую половину личиночного развития головастиков. Не оказывая влияния на динамику численности, эти хищники не могут не оказывать селективного воздействия на темпы онтогенеза головастиков: чем продолжительнее личиночное развитие, тем выше вероятность уничтожения головастика хищником. Следовательно, в каждом поколении хищники должны интенсивно выедать более медленно развивающихся головастиков, определяя тем самым отбор на ускорение темпов онтогенеза. Поскольку более быстрому онтогенезу препятствует эффект группы, отбор хищниками равнозначен отбору на снижение чувствительности головастиков к этому эффекту. Однако этому направлению отбора противодействует ряд факторов, совокупно обусловливающих отбор на поддержание эффекта группы.
Во-первых, это неблагоприятные абиотические условия, т. е. случайно действующие факторы, губительные для выходящих из водоема сеголеток: резкая засуха, сильные дожди, не говоря уже о граде. Э. М. Смирина (1980) отмечала массовую гибель лягушат в результате грозового ливня. Растягивание сроков метаморфоза предохраняет генерацию от массовой гибели под действием подобных случайных факторов. Во-вторых, это животные, питающиеся лягушатами. На пике выхода сеголетки служат кормом паукам-серебрянкам, тритонам, зеленым лягушкам (до 20 сеголеток в желудке), поползням, сойкам, воронам, бурозубкам (до 30 сеголеток в час) - словом, всем, для кого они являются доступной добычей. Естественно, чем выше плотность сеголеток, тем выше пресс хищников. И в этом случае растягивание сроков метаморфоза адаптивно.
Наконец, учеты численности сеголеток при помощи мечения с повторным отловом, проведенные С. М. Ляпковым, показали, вопреки мнению Д. Калли и К. Гравуа (Culley, Gravois, 1972), что существует гибель сеголеток, зависящая от их плотности. На рис. 9 изображены динамика выхода сеголеток из водоема, их численность на учетной площадке (площадь 5300 м2) и смертность. Пик смертности, несколько сдвинутый во времени относительно пика численности, и есть результат ухудшения условий при увеличении плотности популяции. Рост смертности в конце периода выхода сеголеток обусловлен снижением жизнеспособности животных, поздно кончающих метаморфоз. Таким образом, отбору на снижение чувствительности к эффекту группы противодействует отбор на его поддержание, т. е. на растягивание сроков метаморфоза генерации.
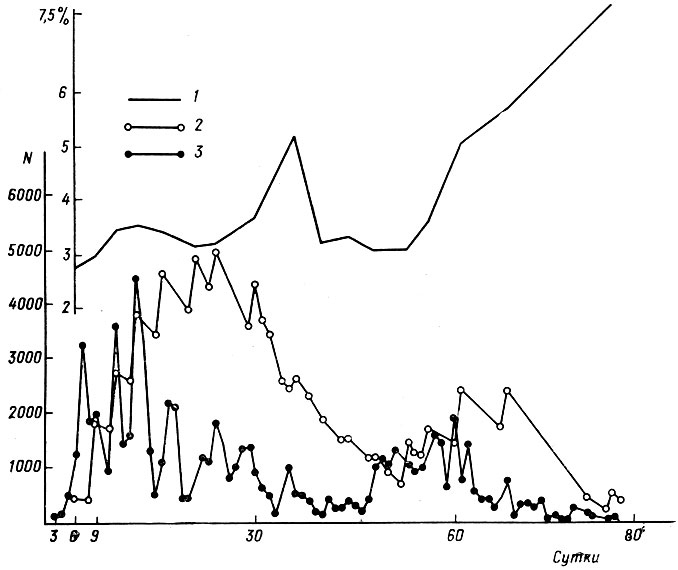
Рис. 9. Динамика выхода из водоема (3), численность на учетной площадке в радиусе 50 м от водоема (2) и смертность сеголеток Rana temporaria (1)
Необходимо подчеркнуть, что увеличение смертности сеголеток, особенно от случайных факторов, неизбежное при сокращении сроков метаморфоза генерации, стимулировало бы не только отбор на поддержание или усиление эффекта группы, но и отбор на плодовитость, компенсирующий случайную гибель. Однако отбор на плодовитость, как показано выше, неэффективен. Ему противодействует зависящая от плотности гибель эмбрионов и головастиков. Иными словами, отбор на плодовитость и противодействующие ему векторы связаны через внутрипопуляционные взаимодействия головастиков с отбором на ускорение онтогенеза и противодействующими ему векторами отбора. В результате эффект группы сохраняется, сохраняется и зависящая от плотности гибель головастиков.
Следовательно, популяция Rana temporaria находится под постоянным воздействием системы векторов движущего отбора, взаимодействующих так, что усиление отбора в любом из рассмотренных направлений автоматически приводит к усилению отбора в противоположном направлении и сохранению популяцией равновесного состояния, т. е. к ее эволюционной стабилизация (рис. 10).
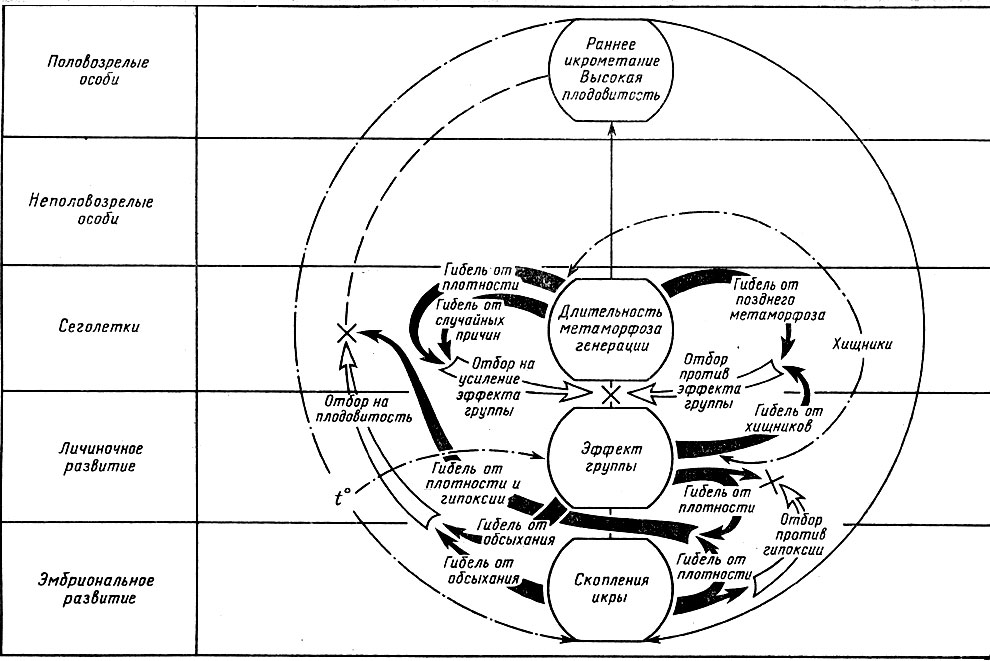
Рис. 10 Схема взаимодействия векторов отбора в популяции Rana temporaria в условиях Подмосковья
Все рассмотренные векторы отбора связаны с признаками, обладающими широкой нормой реакции и поэтому низкой, если не нулевой, наследуемостью. Так как отбор по подобным признакам неэффективен, может возникнуть сомнение в том, что взаимодействие векторов отбора, идущего по этим признакам, стабилизирует популяцию. Однако эффект генокопирования доказан и теоретически (Шмальгаузен, 1968а), и экспериментально (Waddington,. 1957). Следовательно, не низкая наследуемость, а именно взаимное уравновешивание противоположно направленных векторов отбора обусловливает эволюционную стабильность популяции.
Здесь не рассмотрен трехлетний период жизни лягушек - от сеголетки до половой зрелости. Однако экология неполовозрелых особей очень сходна с экологией взрослых, за исключением короткого, занимающего около 10 дней пер'иода икрометания. Во всяком случае, пирамида возрастов лягушек (Ищенко, Леденцов, 1985; Мина, 1974) указывает на равномерность убыли численности генерации. Иными словами, этот период не вносит изменений в предлагаемую модель эволюционной стабильности популяции Rana temporaria. Подтверждение стабилизирующего действия разнонаправленных векторов отбора получено Р. А. Гринько (1987) при изучении механизмов стабилизации плодовитости в лабораторных популяциях Drosophila melanogaster. При сведении к минимуму и стабилизации средовых воздействий плодовитость исследованных популяций в течение 20 поколений слабо флуктуирует вблизи средней. Эта стабильность популяционной плодовитости обусловлена взаимодействием двух спонтанно существующих направлений отбора: отбором в пользу малоплодовитых самок на ранних стадиях онтогенеза и отбором в пользу преимущественного размножения высокоплодовитых мух. Средние по плодовитости особи занимают промежуточное положение. Малоплодовитые самки составляют большую часть популяции и характеризуются повышенным энергетическим обменом, что, видимо, и обусловливает их успех на ранних стадиях онтогенеза и в то же время проигрыш по участию в размножении.
Равновесие векторов отбора, стабилизирующее популяцию, можно было бы считать редким, даже уникальным случаем, если бы ряд примеров сбалансированного полиморфизма и устойчивости отдельных признаков не поддавался интерпретации с вышеизложенных позиций - как результат действия уравновешивающих друг друга направлений отбора. Например, у арктического поморника, по данным О'Дональда (O'Donald, 1972), существуют две основные формы: темно- и светлоокрашенная. Темноокрашенные самцы имеют преимущество при спаривании с любыми самками. Однако светлоокрашенные самки созревают раньше темноокрашенных. В результате в популяции сохраняется постоянная концентрация обеих форм. Сходная ситуация наблюдается у Donaus chrisippus в Восточной Африке. Две морфы этого вида, dorippus и chrisippus, находятся в равновесии, так как самцы chrisippus имеют в течение 3-4 месяцев преимущество в спаривании, а самки dorippus более привлекательны для самцов обеих морф (Smith, 1975). К явлениям того же порядка следует, видимо, отнести полиморфизм по типам поведения у петухов на птицефермах (Кайданов, 1966). Петухи сильного типа поведения имеют преимущество в размножении, пока их мало. Когда их становится больше, они теряют силы и время на драки и установление иерархии, а преимущество в размножении получают петухи слабого типа.
Столь же четко противодействие векторов отбора прослеживается при циклическом отборе у Adalia bipunctata (Тимофеев-Ресовский, Свирежев, 1966). У этих коровок зимой выживает преимущественно темноокрашенная форма, а летом, из-за более высокой плодовитости, преимущество получают красные жуки. Для одного из видов дарвиновых вьюрков - Geospiza fortis - на о-ве Большая Дафна показано (Price, 1984; Price et al., 1984) равновесие двух противоположных направлений отбора: на увеличение высоты клюва и размеров тела, связанное с питанием крупными семенами во время засух, и на уменьшение обоих этих признаков в периоды нормальной влажности. Подобные данные получены и на Geospisa conirostris (Grant, 1985). Аналогично, согласно Д. Лэку (Lack, 1954), размеры кладки у воробьиных представляют собой результат взаимодействия отбора на плодовитость и противодействующего ему отбора на уменьшение количества птенцов, обусловленного трудностью выкармливания большого выводка.
Таким образом, не только популяция в целом, как это показано выше на Rana temporaria, но и отдельные признаки, причем у разных групп животных, стабилизируются противодействующими друг другу векторами отбора. Можно думать, что многие случаи, интерпретируемые ныне как частотно- и плотностно-зависимый отбор, удается лучше понять, если интерпретировать их как взаимодействие разнонаправленных векторов движущего отбора. Иными словами, можно ожидать, что этот механизм эволюционной стабилизации достаточно распространен в природе.
В заключение необходимо подчеркнуть, что представление о равновесии векторов движущего отбора как механизме эволюционной стабилизации популяций диких видов не противоречит теории стабилизирующего отбора, а, напротив, дополняет ее. Именно стабилизация состояния популяции в непрерывно колеблющихся условиях среды и создает базу для действия стабилизирующего отбора как механизма сохранения адаптивных признаков и эволюции процессов онтогенеза.
Как сказано выше, стабилизирующий отбор действует в стабильных условиях среды через элиминацию генотипических уклонений, выходящих за пределы эволюционной нормы (Шмальгаузен, 1968а). Второй случай, при котором может действовать стабилизирующий отбор, - ухудшение условий среды при отборе по стабильным признакам фенотипа. Этот случай иллюстрирует гибель крайних вариантов изменчивости количественных признаков у воробьев (Bumpus, 1899), который сам И. И. Шмальгаузен
(1968) приводил в качестве примера стабилизирующего отбора. При ухудшении условий среды, действующих на пластичные признаки фенотипа, смоделировать стабилизирующий отбор не удается (см. выше). Стабилизирующий отбор эффективен при условии либо стабильности среды, либо стабильности признаков, по которым оп идет. Именно эту ситуацию, т. е. стабилизацию соотношения популяции и среды ее обитания, и создает равновесие векторов движущего отбора, обеспечивая тем самым эффективность отбора стабилизирующего, в первую очередь канализирующей его субформы.
Предлагаемое в данной работе объяснение эволюционной стабилизации популяций диких видов в методическом отношении берет начало от введенной Р. Перлем (Pearl, 1927; Pearl, Parker, 1921) и получившей широкое распространение методики составления таблиц выживания, от которой оно отличается тем, что были проанализированы не только факторы, вызывающие смертность, но и реакции популяции на эти факторы, что и позволило выявить направления отбора. В теоретическом отношении высказанная концепция близка к принципу Холдэна - Семевского. Вышеизложенные данные являются помимо всего остального прямым подтверждением этого принципа, т. е. прямо показывают, что повышение приспособленности в одном каком-либо направлении ведет к снижению приспособленности в других направлениях. Так, "выигрыш" в терморегуляции эмбрионального развития, обусловленный свойственным Rana temporaria способом икрометания (крупные кладки, слипающиеся в сплошные "подушки"), обусловил "проигрыш" в обеспеченности эмбрионального развития кислородом. Аналогично терморегуляция в скоплениях головастиков повлекла за собой усиление неблагоприятных последствий эффекта группы - замедление темпов личиночного развития. Адаптация к икрометанию в более ранние сроки повлекла массовую гибель икры от обсыхания и т. д.
Однако необходимо еще раз подчеркнуть, что в отличие от ситуации, ожидаемой исходя из принципа Холдэна - Семевского, отбор не приводит к эволюционной компенсации снижения приспособленности. В этом отношении он неэффективен или эффективен в очень слабой степени. Тем самым оптимизация соотношения популяции со средой оказывается недостижимой не только экологически, но и эволюционно.
Если сказанное справедливо, то стабилизирующий отбор оказывается причиной и механизмом сохранения не тех признаков, приспособленность которых оптимальна (подобных признаков, по всей вероятности, вообще не существует), а признаков, приспособленность которых не может быть изменена в данных условиях среды. Другими словами, стабилизирующий отбор фиксирует несовершенные с точки зрения их приспособительного значения дефинитивные признаки, а затем совершенствует процессы онтогенеза подобных неоптимальных адаптаций. А. М. Уголев (1961, 1985) специально подчеркивал несовершенство физиологических процессов у высших организмов. Из этого следует, что формирующийся главным образом посредством стабилизирующего отбора тип онтогенеза, а тем самым и структурный тип (тип организации) надвидовых таксонов представляет собой результат оптимизации морфогенеза неоптимальных дефинитивных структур. Следовательно, уже сформировавшаяся структура, являющаяся важнейшим параметром, определяющим направление филогенеза (см. раздел II), перестраивается в ходе этого процесса не только благодаря изменениям, вызванным изменениями среды, но и в силу необходимости дальнейшей адаптации ранее возникших приспособлений.
Представление о том, что эволюционная стабильность популяций определяется равновесием действующих на нее векторов отбора, ставит вопрос о границах устойчивости подобного равновесия. Вероятно, покуда вид существует, несмотря на различия в условиях его существования в разных частях ареала, для каждой из популяций это равновесие выдерживается в целом, невзирая на различия в интенсивности тех или иных векторов отбора.
Косвенным подтверждением этому могут служить многолетние (1968-1974 гг.) исследования И. А. Шилова с сотрудниками (Шилов и др., 1973а, б; 1975) по экологии полевки-экономки (Microtus oeconomus Pall.) в Дарвинском государственном заповеднике на берегу и островах Рыбинского водохранилища. Это водохранилище начало наполняться в 1941 г., и формирование его завершилось в 1947 г. Заполнение водохранилища внесло глубокие изменения в экологию популяций полевки-экономки, обитавшей по берегам рек Шексны и Мологи, междуречье которых было занято водохранилищем. Зарегулирование стока рек в сочетании с пологими берегами привело к изменению паводкового режима. Подъем воды в водохранилище начинается со снеготаянием, но спадает вода очень медленно, из-за чего к октябрю уровень воды все еще выше, чем весной, перед началом ее подъема. Поэтому прибрежная популяция "пульсирует". При подъеме воды, пик которого приходится на май - июнь, она оттесняется к верхней границе зоны временного затопления, а по мере ее спада и разрастания околоводной растительности распространяется по этой зоне вслед за отступающей водой. Ширина зоны в зависимости от крутизны берегов составляет от нескольких метров до 5-6 км. Кроме того, существуют группировки полевок, занимающие подходящие для них стации в верховьях ручьев, не подверженных влиянию колебаний уровня водохранилища, т. е. обитающие в условиях, более сходных с типичными для вида.
После заполнения чаши водохранилища часть отложений торфа затопленных сфагновых болот оторвалась от дна и всплыла, образовав плавучие, но не дрейфующие острова. "Большой торфяник", на котором проводились работы, представляет собой остров длиной 12 км и шириной от 4 км до нескольких сот метров. Во время подъема уровня водохранилища он не затапливается. К началу работ остров был вновь заселен высшими растениями, включая кусты ивы и молодые осинки. Полевка-экономка заселила его, по-видимому, примерно за 25 лет до начала работ, попав сюда скорее всего с плавающих островков. Изоляция островной популяции полная и поддерживается таковой около 50 поколений. До ближайшей береговой популяции расстояние 10 км.
Авторы исследовали динамику численности, половую, возрастную и пространственную структуры популяций, степень стрессированности особей, входящих в популяционные группировки, и ряд морфологических признаков, характеризующих физиологическое состояние отловленных зверьков. Кроме массы и длины тела определяли: относительные массы почки (%), межлопаточного бурого жира (%), надпочечников (%), тимуса (%), число эритроцитов (млн в 1 мм3) и гемоглобин (мг/%). Окраску меха оценивали в процентах отраженного света.
Различия в экологии береговой и островной популяций довольно значительны. Как уже говорилось, береговая "пульсирует", а островная стабильна по территории. Различна динамика численности, главным образом в соотношении численностей осенью и весной. На торфяном острове полевки лишь иногда делают норы в высоких кочках, а обычно живут в гнездах на поверхности торфа, так как вода стоит непосредственно под поверхностью. Весенняя численность (перезимовавшие зверьки) в береговой популяции относительно выше, чем в популяции, живущей на острове, и колебания численности береговой популяции выражены резче. Однако в морфофизиологических показателях различия практически отсутствуют. У полевок, отловленных на острове, относительная масса почки несколько выше, а относительная масса бурого жира несколько меньше, что объясняется более суровым микроклиматом торфов: постоянными ветрами и высокой влажностью приземного слоя воздуха. Остальные признаки не различаются. Таким образом, различия в экологии существенно более выражены, чем в физиологии.
Прежде всего, вслед за И. А. Шиловым необходимо подчеркнуть, что 50 поколений существования изолированной островной популяции не привели к сколь-нибудь заметной ее дивергенции от береговой. Объяснить отсутствие дивергенции выбором полевками каких-то микростаций, обеспечивающих возможность их существования в одинаковых условиях, не удается. Различия в структуре поселений, динамике численности, гнездостроении свидетельствуют об обратном. Можно было бы предположить, что различия в условиях существования находятся в пределах нормы реакции исследованных, заведомо пластичных признаков фенотипа. Тогда различия между популяциями должны были бы быть более заметными, во всяком случае затрагивать большее число признаков, характеризующих энергетику, чем это наблюдается в действительности. Иными словами, за исключением двух вышеуказанных признаков, нет фенотипической адаптации к новым условиям. Объяснить это можно, по-видимому, только тем, что, несмотря на изменение динамики численности, которое указывает на изменение интенсивности отбора по сезонам года, общее равновесие векторов отбора не нарушено, значит, изменения в экологии оказались недостаточными для смещения равновесия и, следовательно, для дивергенции популяций.
Таким образом, литературные данные позволяют предположить, что механизм эволюционной стабилизации популяций на основе взаимного уравновешивания действующих в них векторсв отбора достаточно распространен. Если это так, то получает объяснение обычная для биологии ситуация, когда эволюция (в смысле новообразования) даже на популяционно-видовом уровне не является непосредственно наблюдаемым процессом и лишь подразумевается в прошлом на основе изучения ныне существующих адаптаций. Лишь в случае действительно резких изменений среды, главным образом антропогенных (пестициды, задымление и т. п.), т. е. действия эволюционно новых факторов, обусловливающих возникновение новых, не уравновешенных векторов отбора, эволюция проявляется непосредственно.
Вероятно, ближайшим эволюционным следствием, обусловленным равновесием действующих на популяцию векторов отбора, является формирование пластичных признаков фенотипа и (или) сбалансированного полиморфизма. Оба эти механизма, а особенно первый из них, защищают популяцию от случайных флуктуаций внешних условий, т. е. сохраняют ее положение в окружающей среде. Выработку и тех и других типов признаков определяет само противодействие векторов отбора, заставляющее организмы одновременно приспосабливаться к альтернативным, в лучшем случае, чередующимся во времени экологическим воздействиям.
С позиций равновесного состояния популяций под действием системы векторов отбора находит объяснение и широкий диапазон изменений темпов эволюции популяций. Изменение тех факторов среды, к которым вид в какой-то мере уже приспособлен, т. е. "эволюционно освоенных факторов", приведет к изменению популяции в новую точку равновесия. Для пластичных признаков фенотипа это равносильно смещению в пределах нормы реакции (с учетом скрытого резерва адаптивного реагирования). Для признаков со сбалансированным полиморфизмом подобное изменение приведет к новому балансу частот. В обоих случаях подобные изменения будут соответствовать существующим представлениям о микроэволюции, включая и представление об "элементарном эволюционном акте" (Тимофеев-Ресовский и др., 1969). Однако популяция не изменит своего видоспецифического облика.
Если подобная эволюция будет происходить достаточно долго и медленно, генокопирование сохранит соответствие нормы реакции или полиморфизма диапазону колебаний среды. В этом случае взаимодействие векторов отбора не нарушится и сохранится тренд эволюции, который независимо от ортоселекции определяет направленность филогенеза (см. гл. 2).
Достаточно резкое, выходящее за пределы реагирования изменение среды приведет к перестройке взаимодействия векторов отбора, что равносильно изменению направления эволюции. Дисбаланс векторов равноценен возникновению нового, несбалансированного другими направления отбора. Иными словами, изменение направления эволюции популяции может быть вызвано не только перестройкой экологической обстановки, но и скоростью этой перестройки. К тем же результатам приведет и возникновение нового вектора отбора при столкновении популяции с "эволюционно не освоенными" факторами среды, например с антропогенными воздействиями. Оба последних случая сводятся к возникновению новой равнодействующей всех векторов отбора, т. е. к одному из следствий принципа Холдэна - Семевского (см. выше). Таким образом, представление об эволюционной стабилизации естественных популяций под действием системы взаимно уравновешивающих друг друга векторов отбора позволяет дополнить существующие представления о причинах изменения направления микроэволюции. Подобные изменения определяются: 1) изменением условий среды, что приводит к изменению направлений отбора; 2) изменением качественного состава наследственной изменчивости, подвергающейся отбору в данных условиях среды; 3) изменением темпов эволюции, не позволяющим популяции сохранить то же направление эволюции из-за перестройки системы влияющих на нее векторов отбора.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'