Глава 2. Направленность макроэволюции
2.1. Соотношение микроэволюции и макроэволюции
Проблема соотношения микроэволюции и макроэволюции неизменно привлекает внимание исследователей, главным образом по двум причинам. Во-первых, эта проблема действительно имеет существенное методологическое значение, так как подразумевает вопрос о единстве механизмов и закономерностей эволюции, т. е. вопрос о структуре эволюционной теории в целом. Во-вторых, обсуждение этой проблемы не требует специальных исследований, т. е. может быть основано лишь на общих соображениях.
Термины "микро-" и "макроэволюция" впервые применил, по-видимому, Ю. А. Филипченко (Philiptschenko, 1927), затем их использовали Г. Добжанский (Dobzhansky, 1937), Р. Гольдшмит (Goldsmidt, 1940), затем Дж. Симпсон (Симпсон, 1948; Simpson, 1944). Перечисленным авторам принадлежит и общеупотребительное в современной литературе разграничение микроэволюции как эволюции популяций, идущей под действием естественного отбора и оканчивающейся видообразованием, и макроэволюции как эволюции надвидовых таксонов. Дж. Симпсон (1948) ввел еще один термин - "мегаэволюция", подразумевая под ним эволюцию таксонов высокого ранга - классов и выше. Термином "макроэволюция" он обозначил эволюцию родов, семейств, отрядов. Это более дробное разграничение уровней эволюции не получило распространения. В то же время следует признать, что именно благодаря Симпсону термины "микро-" и "макроэволюция" вошли в биологическую терминологию (Галл, 1983; Грант, 1980). Дж. Симпсон (1948; Simpson, 1953) придерживался мнения, что и микро-, и макро-, и мегаэволюция происходят под действием дарвиновских факторов эволюции - наследственной изменчивости и отбора.
Противоположная точка зрения, согласно которой таксоны надвидового ранга возникают скачкообразно, восходит к Л. Кено (Cuenot, 1914) и Л. С. Бергу (1922). Ю. А. Филипченко (1927) также считал, что макроэволюция происходит посредством макромутаций. Р. Гольдшмит (Goldsmidt, 1940) придерживался этой же точки зрения, предполагая, так же как и Л. Кено, что макромутации создают формы, преадаптированные к тем средам, в которых обитают систематические группы - потомки подобных "многообещающих уродов". Представления о макрогенезе получили поддержку и среди палеонтологов (Красилов, 1980; Личков, 1965; Schindewolf, 1952, 1963). Т. Девилер (Devillers, 1965) предположил, что возникновение надвидовых таксонов связано с мутационными изменениями морфогенеза. В его экспериментах экстирпация или подсадка окелетогенной мезенхимы в область зачатка конечности позвоночных приводила соответственно к недоразвитию или удвоению костей голени. Например, у птиц развивалась малая берцовая кость - признак класса. В новейших обоснованиях макрогенеза используется представление о латеральном переносе генов (Кордюм, 1982). Таким образом, представления о собственном, сальтационном механизме макроэволюции до сих пор имеют сторонников.
При интерпретации отдельных мутаций или нарушений формообразования как механизма сальтационного возникновения надвидовых таксонов обычно не обращают внимания на то, что измененные признаки не являются эволюционно новыми. В качестве таксономически значимых они интерпретируются только по аналогии с признаками уже существующих систематических групп. Например, мутация дрозофилы tetraptera может быть интерпретирована как изменение диагностического признака отряда Diptera только потому, что существуют четверокрылые насекомые, от которых возникли двукрылые. Удвоение костей голени птиц можно интерпретировать как меняющее признак этого класса только на том основании, что предки птиц - рептилии - имели две берцовые кости. Напротив, мутация дрозофилы aristopedia, несмотря на все ее своеобразие - ножка вместо аристы, не может иметь таксономической интерпретации, так как среди насекомых нет форм с ногами на голове. Иными словами, представления о сальтационном механизме макроэволюции основаны на порочном логическом круге. В качестве экспериментального обоснования этих представлений используются искусственно вызванные атавизмы, а не новые адаптации. В случае новых адаптаций им нельзя было бы приписать тот или иной конкретный таксономический ранг.
Наиболее глубокий и в настоящее время общепризнанный анализ причин того, что вся эволюция идет на основе комплексов малых мутаций, принадлежит И. И. Шмальгаузену (1968а). На уровне молекулярной генетики, в том числе применительно и к латеральному переносу генов, аналогичный анализ дан Р. В. Хейсиным (1984). Обсуждение этих концепций выходит за рамки работы. Единство механизмов микро- и макроэволюции подчеркивали очень многие авторы (Галл, 1980, 1983; Майр, 1968; Мина, 1986; Шварц, 1969; Воск, 1965; Hecht, 1965; Rencsh, I960; Simpson, 1953; Van Valen, 1963 и др.). В настоящее время эта точка зрения, несомненно, господствует, что показали специально посвященная соотношению микро- и макроэволюции конференция в Тарту (1980) и дискуссия на эту тему, проведенная во время международной конференции "Эволюция и среда" (ЧССР, Брно, 1981) (Смирнов, 1980; Тимофеев-Ресовский, 1980; Я блоков, 1980).
В то же время большинство авторов, обсуждающих соотношение этих двух уровней рассмотрения эволюционного процесса, подчеркивают несводимость макроэволюции к микроэволюции (Паавер, 1980; Тимофеев-Ресовский, 1980; Яблоков, 1980; Воск, 1965; Dobzhansky, 1951; Hecht, 1965). А. П. Расницин (1969) и В. В. Жерихин (Жерихин, Расницын, 1980) подчеркнули несоответствие темпов макроэволюции тем, которые можно было бы ожидать на основе теории микроэволюции. С. С. Шварц (1969) писал об организационной и экологической предопределенности макроэволюции, Н. В. Тимофеев-Ресовский (1980)) и И. Элдредж (Eldredge, 1986) сослались на "межвидовой отбор".
Получается, что вопрос о своеобразии механизмов микро- и макроэволюции в настоящее время утратил актуальность, тогда как вопрос о сводимости или несводимости макроэволюции к микроэволюции продолжает обсуждаться. Своеобразие микро- и макроэволюции очевидно, поэтому целесообразно обсуждать не само своеобразие, а его причины и этим путем попытаться выяснить вопрос о сводимости или несводимости закономерностей макроэволюции к микроэволюционным закономерностям.
Обсуждая этот вопрос, необходимо напомнить, что с 60-х гг. прошлого века процесс эволюции надвидовых таксонов обозначался термином "филогенез". Автор этого термина Э. Геккель (Haechel, 1866, 1894) и определил филогенез как историческое развитие систематических групп. В такой интерпретации филогенез и макроэволюция - синонимы. Однако затем под филогенезом стали понимать не только историческое развитие таксонов, но и историческое развитие отдельных признаков организма и организма как целого. Строгое определение филогенеза как исторического ряда известных (т. е. прошедших естественный отбор) отногенезов дал И. И. Шмальгаузен (1969). При использовании этого определения соотношение микроэволюции и филогенеза описывается концепцией сетчатого родства особей в популяции (Шмальгаузен, 1969), объясняющей соотношение преемственности онтогенезов внутри эволюционирующей популяции и преемственность таксонов в ходе их исторического развития.
Отношения особей, составляющих популяцию, можно представить трехмерной решеткой, в которой каждый узел - скрещивание прошедших отбор особей, а каждая линия - онтогенез выживших, т. е. прошедших отбор, потомков этих особей. Популяция же в целом, первоначально принадлежавшая исходному "родительскому" таксону, в том случае, если она эволюционирует в смысле преобразования организации составляющих ее особей, в конце концов становится одной из популяций дочернего таксона. В этой модели ретроспективный анализ преобразований организации от конечного состояния популяции к исходному строго соответствует данному И. И. Шмальгаузеном (1969) определению филогенеза. Теоретически, прослеживая по ячейкам решетки преобразования организации от потомков к предкам, можно было бы проследить исторический ряд онтогенезов от любого представителя дочернего таксона до того представителя исходного таксона, у которого тенденция к эволюции в сторону дочернего таксона впервые проявляется как индивидуальное уклонение.
На практике подобная реконструкция филогенеза невозможна. Во-первых, технически невозможно настолько полно исследовать организацию, а тем более онтогенез особей, чтобы судить о филогенезе как об историческом ряде онтогенезов. Во-вторых, неполнота палеонтологической летописи не позволяет, за очень редкими исключениями, подобрать исторический ряд форм, связанных между собой непрерывной преемственностью поколений. Мало того, в тех редких случаях, когда в руках палеонтологов находится материал, позволяющий анализировать индивидуальную изменчивость в череде поколений, такой как Paludina neimajri или Planorbis multiformis, в точном соответствии с концепцией сетчатого родства невозможно выделить единственный филогенетический ряд, отражающий исторические преобразования популяции. Напротив, внутри популяции легко выделить множество равноценных друг другу подобных рядов, каждый из которых характеризует исторические преобразования популяции и филогенеза таксонов.
Поскольку достаточно полное описание исторических изменений организации особи, а тем более всего онтогенеза невозможно, реальные реконструкции филогенеза основаны на изучении исторического развития отдельных признаков. Основы методологии реконструкции филогенеза признаков созданы Э. Геккелем (Haeckel, 1866) в форме метода тройного параллелизма, который видоизменялся и совершенствовался по мере накопления опыта филогенетических исследований. Относительно методологии реконструкции филогенеза существует обширная литература, возникшая благодаря работам В. Хеннига (Hennig, 1966). Обзоры этой литературы даны А. П. Расницыным, Г. М.Длусеким (1988) и Л. П. Татариновым (1987а, б). Обсуждение трудностей и эффективности выявления апоморфных и плезиоморфных признаков выходит за пределы задачи данной книги. Реконструкции филогенеза отдельных признаков затем экстраполируются на филогенез соответствующего таксона. В настоящее время, когда систематика и историческое развитие большей части вымерших и современных систематических групп в ранге отрядов - классов более или менее известны, данные по филогенезу признаков служат для уточнения или перестройки существующих реконструкций филогенеза таксонов. Для плохо изученных групп ряды по признаку (признакам) непосредственно служат реконструкциями филогенеза таксонов: последовательность преобразований признака условно отождествляется с последовательностью исторического развития систематической группы. Ранг таксона, филогенез которого реконструируется посредством подобной экстраполяции, зависит от признаков, используемых для построения ряда. Изучение филогенеза таксономических признаков класса, например класса млекопитающих или класса амфибий, позволяет судить об историческом развитии именно этих таксонов. Таким образом, при реконструкции филогенеза надвидовых таксонов, тождественной реконструкции макроэволюций этих таксонов, производится экстраполяция реконструкций филогенеза отдельных признаков на макроэволюции систематической группы соответствующего ранга.
На самом деле экстраполяция выводов о филогенезе признаков на таксон подразумевает два этапа: с признаков на филогенез в строгом смысле термина - исторический ряд онтогенезов - и затем уже на таксон соответствующего ранга. Подобная двойная экстраполяция более точно соответствовала бы концепции сетчатого родства особей в популяции, однако в силу вышеизложенных причин она невозможна, и промежуточный этап лишь подразумевается. Любая экстраполяция с частного на общее и экстраполяция, осуществляемая при филогенетических исследованиях в том числе, содержит в себе возможность ошибок. Часть подобных ошибок обусловлена несовершенством филогенетических (на самом деле - сравнительных) рядов по признакам, часть - неправомочностью экстраполяции с признака на таксон. Оба типа ошибок могут быть вызваны несколькими причинами: техническими затруднениями - недостатком материала (не только палеонтологического); сложностями, обусловленными неравномерностью темпов эволюции подсистем организма (гетеробатмией), и перекрестом специализации, что приводит к включению в один ряд признаков параллельных филумов; неадекватным задаче подбором признаков и другими причинами. Попытки интерпретировать исторические преобразования даже морфологических признаков как адаптивные, что при успехе, несомненно, повышает надежность филогенетических реконструкций (Юдин, 1970, 1974), часто затруднены, поскольку мультифункциональность признаков зачастую не поддается достаточно полному анализу. Кроме того, многие признаки, используемые в филогенетических исследованиях, представляют собой координационные преобразования компонентов функциональных систем, т. е. имеют лишь косвенное адаптивное значение. Перечисленные причины ошибок, а приведенный их список далеко не полон, обусловливают необходимость дальнейших ревизий существующих филогенетических реконструкций. Поиски же объективных подходов в филогенетических исследованиях породили проблему таксономически и филогенетически значимых признаков (Лебедкина, 1980).
Описанная процедура реконструкции филогенеза систематических групп представляет собой, таким образом, процесс постепенного приближения эмпирических реконструкций к теоретической модели, описываемой концепцией сетчатого родства. Поскольку полное совпадение, видимо, никогда не может быть достигнуто, филогенетические схемы никогда не могут быть абсолютно точными. Сказанное, однако, не означает, что филогенетические реконструкции всегда ненадежны, напротив, как отмечал К. А. Юдин (1974), происхождение групп в ранге подкласса - надкласса установлено во многих случаях вполне надежно. Однако филогенез таксонов более низкого ранга, или, что то же самое, детализация существующих реконструкций (некоторое исключение может составлять видовой уровень), а также реконструкции происхождения типов постоянно служат предметом дискуссий и обычно далеки от однозначной интерпретации.
Из сказанного о методах реконструкции филогенеза таксонов, т. е. макроэволюции, следует, что своеобразие закономерностей этого процесса обусловлено изменением объекта изучения и масштаба времени, в течение которого прослеживается эволюция. Исследователь, занимающийся микроэволюцией, имеет дело в первую очередь с индивидуальной изменчивостью и ее преобразованиями под действием факторов эволюции. Согласно замечанию Р. Гольдшмита (Goldsmidt, 1940), эти изменения по своей продолжительности сравнимы с продолжительностью человеческой жизни. Напротив, исследователь, занимающийся макроэволюцией, имеет дело с "геологическими" промежутками времени, от десятков тысяч до десятков и сотен миллионов лет. Объект его непосредственного изучения - исторические преобразования отдельных признаков или их комплексов на всех уровнях организации особи, от макроморфологического до молекулярного. Эти преобразования прямо или косвенно адаптивны, т. е. представляют собой следствия действия отбора в популяциях. Различия в объекте и связанные с ними различия в подходах и методах исследования неизбежно определяют своеобразие закономерностей микро- и макроэволюции.
То, что микро- и макроэволюция лишь разные аспекты единого процесса, очевидно для большинства авторов (см. выше). Однако это еще не решение вопроса о сводимости или несводимости макроэволюционного процесса к микроэволюционному. С позиций вышеизложенной концепции сетчатого родства особей в популяции этот вопрос принимает несколько иную окраску. Речь идет не о редукции (в методологическом значении этого термина) закономерностей высшего порядка (макроэволюционных) к таковым низшего порядка (микроэволюционным), а о том, что преобразования популяции под действием отбора являются причиной исторических преобразований организации и тем самым макроэволюции. Следствие - в данном случае макроэволюцию можно объяснить причиной - микроэволюцией, но сводить следствие к причине не представляется возможным. Между ними можно только установить соответствие: это соответствие причины и следствия, т. е. направление отбора в популяциях и преобразований организации в филогенезе часто неоднозначно. Этим, еще больше чем изменением объекта, объясняется своеобразие закономерностей макроэволюции. Неоднозначность причинно-следственных связей между микро- и макроэволюцией обусловлена, по-видимому, двумя основными причинами: сложностью, точнее, системностью организации особи и нелинейностью по времени изменений ее организации, что наблюдается даже при длительном сохранении направления действия соответствующего вектора отбора.
Так, например, и хвостатые, и бесхвостые амфибии ловят добычу путем выбрасывания языка (рис. 11). Направление отбора - совершенствование ловли добычи и механизма выбрасывания языка - едино. Но у хвостатых амфибий язьж сохранил первичную для Tetrapoda связь с подъязычно-жаберным скелетом, и совершенствование механизма его движений в ряду Urodela основано на высвобождении базального элемента (копулы) от связи с гиоидной дугой (рис. 12-15). У взрослых бесхвостых амфибий в результате перестройки морфологии подъязычно-жаберного скелета на стадии головастика связь языка с подъязычно- жаберным скелетом была утрачена (рис. 16). Это привело к формированию гидравлического механизма выбрасывания языка. Причем среди Апига дальнейшее совершенствование этого механизма также осуществляется разными способами.
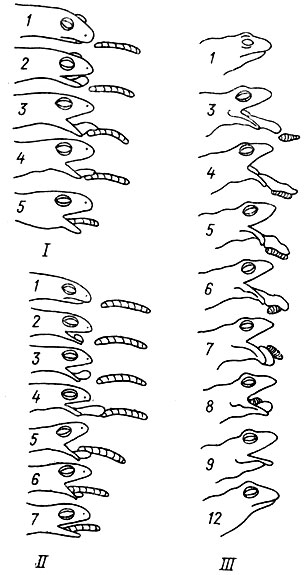
Рис. 11 Выбрасывание языка Ranodoti sibiricus (I), cristatus (II) и Rana temporaria (III). Прорисовка по кинокадрам. Арабскими цифрами обозначены номера кинкадров
Таким образом, у двух отрядов амфибий в силу разной конструктивной основы возникли два разных ответа на одно и то же направление отбора - два разных следствия одной и той же причины. Это очень грубый пример, в котором далеко не полно рассмотрены лишь функциональные, но не онтогенетические и филогенетические связи. Реальные связи, даже функциональные, гораздо сложнее. Однако й этот пример показывает, что следствие, т. е. макроэволюция, несводимо к ее причине - микроэволюции.
По-видимому, первым понял специфику филогенеза как следствия макроэволюции и в полной мере использовал в своих работах это важное в методологическом отношении соподчинение А. Н. Северцов (1939). Во введении к "Морфологическим закономерностям эволюции" он сформулировал две основные задачи разработки проблем филогенеза: "1. Изучение хода эволюции и ее закономерностей и 2. Изучение причин филогенетических изменений организмов" (с. 80). При этом А. Н. Северцов подчеркнул, что на начало 30-х гг. XX в., когда он заканчивал работу над цитируемой книгой, "...хуже обстоит дело со второй задачей дарвинизма, а именно с причинным объяснением эволюции. Выше мы отметили, что, несмотря на блестящие успехи теории наследственности, результаты генетических исследований мало применялись для решения эволюционных вопросов" (с. 80). Под выяснением причин филогенетических изменений организмов А. Н. Северцов понимал прежде всего онтогенетические и функциональные причины, которые он и рассматривал как следствие действия факторов эволюции. По поводу причин функциональных изменений органов он писал: "Для данного вида безразлично, выполняется ли данная функция у потомков тем же органом, что и у предков, или же она выполняется каким-либо другим органом, лишь бы биологически, т. е. с точки зрения выживания в борьбе за существование она выполнялась лучше, чем у предков" (с. 345).
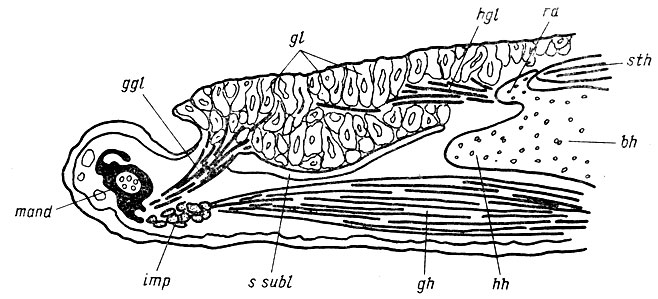
Рис. 12. Язык Hynosius keyscrlingii, схема по сагиттальному срезу дна ротоглоточной полости Основную массу языка образуют многоклеточные слизистые железы (gl), открывающиеся на его спинке. Протракция языка достигается смещением вперед копулы (bh), тесно связанной с языком radii ant. (rа) (см. рис. 13). Собственные мышцы языка, m. genioglossus (ggl) и m. hyoglossus (hgl) обеспечивают изменения конфигурации поверхности языка, что способствует более надежному приклеиванию добычи. Сама возможность протракции языка за пределы рта, ограниченного нижней челюстью (mand), обусловлена наличием лимфатической полости sinus sublingualis (5. subl), которая обособливает язык от подстилающих его структур
По поводу филэмбриогенезов, конкретно применительно к архаллаксисам и ценогенезам, А. Н. Северцов писал: "Нерегулирующиеся эмбриональные вариации, имеющие место в начальных стадиях развития, дают материал для эволюции признаков взрослых животных, происходящей путем архаллаксиса...", и далее: "Не входя в подробное обсуждение вопроса о происождении ценогенезов, отметим все же, что все эти образования происходят, по нашему мнению, путем суммирования естественным отбором регулирующихся эмбриональных вариаций" (с. 520). Из вышеизложенных цитат достаточно ясно, что А. Н. Северцов последовательно придерживался представления о том, что первопричиной филогенетических изменений являются дарвиновские факторы эволюции.
Таким образом, для А. Н. Северцова проблемы соотношения микро- и макроэволюции не существовало, для него соотношение дарвиновских движущих сил эволюции, действующих на популяционном (внутривидовом) уровне, и филогенетического развития надвидовых таксонов было очевидно. Очевидно было оно и для И. И. Шмальгаузена, который не пользовался терминами "микро-" и "макроэволюция", хотя в то время, когда он работал над "Факторами эволюции" (1-е изд. в 1946 г.), эти термины уже широко применялись. Подход И. И. Шмальгаузена к соотношению эволюции на популяционном и надвидовом уровне был тот же, что и у А. Н. Северцова. Я. М. Галл (1980, 1983) подчеркнул, что И. И. Шмальгаузен и Дж. Хаксли внесли кардинальный вклад в разработку проблем соотношения микро- и макроэволюции, объяснив ряд макроэволюционных закономерностей с позиций микроэволюции. Подобное объяснение возможно только при ясном осознании причинно-следственного характера связей между обоими уровнями рассмотрения эволюционного процесса. Иными словами, и для этих авторов проблемы соотношения микро- и макроэволюции тоже не существовало.
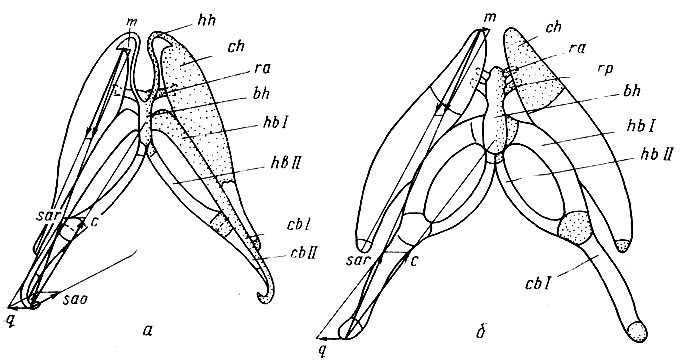
Рис. 13. Подъязычный скелет Ranodon sibiricus (Hynobiidae - а) и Triturus cristatus (Salamandridae - б). Вид с вентральной стороны Слева на каждом рисунке изображены векторы сил, возникающих при сокращении мышц протрагирующих задние рога и копулу, что приводит к выбрасыванию языка. У R. sibiricus в этом движении участвуют две мышцы: m. subarcualis rect. (sar) и m. subarcualis obi. (sao), у Tr. cristatus лишь m. subarcualis rect. (sar). Сила этих мышц (с) приложена к каудальным концам задних рогов, образованным ceratobranchialia I (cb I), и передается через все элементы, образующие эти рога: ceratobranchiale II (cb II) и hypobranchialia I, II (hb I, hb II) на копулу (bh). Передние рога (ch) от каудалыюго смещения удерживают связки, соединяющие задний конец каждого из них, так, что их передние концы испытывают лишь слабое медиальное смещение (m). Латеральному смещению каудальных концов задних рогов (q) препятствуют жесткость их конструкции и сокращение m. subaracualis obi. у R. sibiricus. Тонкие hypohyalia (hh), соединяющие копулу с передними рогами у Hynobiidae не препятствуют протракции языка, связанного с radii ant. (rа), но затрудняют ее, у Salamandridae они редуцируются, превращаясь в radii post, (rр), дополнительно укрепляющие связь языка с копулой
Поэтому возникает вопрос, как и почему проблема соотношения микро- и макроэволюции стала проблемой. Как показано выше, термины "микроэволюция" и "макроэволюция" возникли в генетике и обсуждались в первую очередь генетиками, т. е. биологами, не занимавшимися специально филогенетическими исследованиями. За 50 с лишним лет, прошедших с того времени, когда А. Н. Северцов работал над завершением "Морфологических закономерностей эволюции", ситуация кардинально изменилась: микроэволюционные исследования, основанные главным образом на данных генетики, продвинулись далеко вперед в причинном объяснении эволюции, тогда как макроэволюционные, филогенетические исследования отступили на второй план. В результате соотношение микро- и макроэволюции продолжали обсуждать исходя из микроэволюционных исследований, что недостаточно для понимания их взаимосвязи. Недостаточным оказался и палеонтологический подход, так как он не объяснял своеобразия закономер-ностей макроэволюции, а позволял лишь (констатировать это своеобразие. Видимо, только комплексный эволюционно-морфологический подход дает возможность увидеть причинно-следственный характер связей между закономерностями микро- и макроэволюции. Макроэволюция действительно несводима к микроэволюции просто в силу того, что следствие не сводится к причине, а макроэволюция представляет собой не только следствие микроэволюции, но и специфический уровень рассмотрения эволюционного процесса. Таким образом, проблема соотношения микроэволюции и макроэволюции возникла по недоразумению и не может считаться реально существующей проблемой теории эволюции.
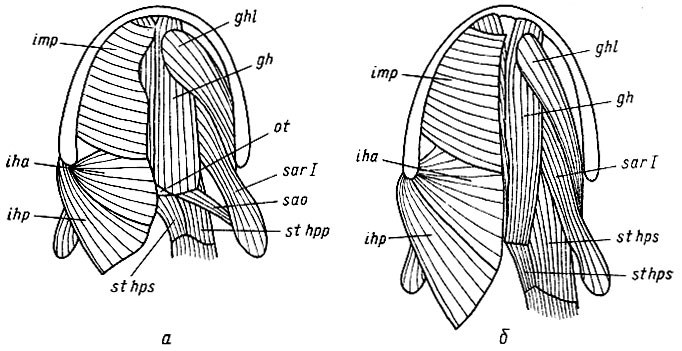
Рис. 14. Мускулатура подъязычного аппарата Ranodon sibiricus (Hynobiidae - а) и Triturus cristatus (Salamandridae - б). Вид с вентральной стороны. Справа удалены поверхностные мышцы: gh - m. geniohyoideus medialis, ghl - m. geniohyoideus lat., iha - m. interhyoideus ant., ihp - m. interhyoideus post., imp - m. intermandibularis post. Сокращение перечисленных мышц поднимает и напрягает дно ротоглоточной полости. Это движение проталкивает воздух в легкие, удаляет его избыток через ноздри и создает опору для горизонтальных движений подъязычного аппарата, обусловливающих протракцию и ретракцию языка, ot - os triquetrum, sao - m. subarcualis obi., sar I - m. subareualis rect. - эти мышцы протрагируют подъязычный аппарат и язык (см. рис. 13). sthpp - pars profundus m. sternohyoideus - эта мышца ретрагирует подъязычный аппарат и язык, sthps - pars superficialis m. sternohyoideus - эта мышца участвует в напряжении дна ротоглоточной полости, создавая опору на os triquetrum для m. geniohyoideus med
Вопрос о соотношении микро- и макроэволюции важен с точки зрения проблемы направленности эволюционного процесса прежде всего потому, что именно изучение макроэволюции всегда служило источником антидарвинистских представлений не только о движущих силах, но и о причинах направленности филогенеза- Собственно, сама проблема направленности эволюции была впервые поставлена Ламарком в его теории градации, движущей силой которой, согласно создателю первой теории эволюции, служит стремление организмов к совершенствованию (см. 2.6). В дальнейшем труды, в первую очередь палеонтологов, породили и поддержали идею ортогенеза, эволюции без дивергенции, в которой представление о направленности филогенеза доведено до абсурда. Этому аспекту критики дарвинизма сторонники его смогли противопоставить лишь две идеи, объясняющие ортогенез признаков и одновременно связывающие теории микро- и макроэволюции. Идею ортоселекции (Plate, 1912) и идею положительной обратной связи, жестко направляющей преобразования признаков под действием отбора (Гиляров, 1970а, б; Татаринов, 1987а, б; Wilson, 1975). Естественно, проблема направленности макроэволюции не исчерпывается этими двумя очень важными положениями. Обсуждая ее, необходимо учитывать и причины направленности филогенеза в целом, и различные аспекты самого явления направленности.
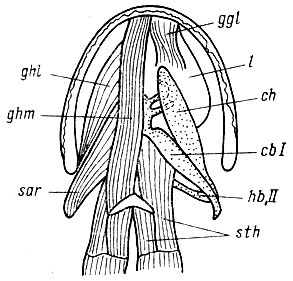
Рис. 15. Подъязычный аппарат безлегочной кавказской саламандры Mertensiella саucasica. Вид с вентральной стороны. Обозначения, как на рис. 12-14. Поверхностные мышцы удалены полностью. Справа на рисунке удалены все остальные мышцы, кроме m. sternohyoideus, для того чтобы показать подъязычный скелет и язык (l). По сравнению с Ranodon sibiricus и Triturus cristatus видно ослабление рогов и прогрессивное развитие копулы, передних лучей, задних лучей и языка, т. е. структур, обеспечивающих захватывание пищи в ущерб структурам, обеспечивающим дыхание
Поскольку проблема направленности макроэволюции имеет длительную историю, богатую не только полемикой, но и обобщениями, определяющими взгляды современных эволюционистов, представляется целесообразным не ограничиваться при ее обсуждении краткими ссылками на литературные источники, а более или менее подробно изложить существующие представления, для того чтобы их критический анализ был возможно более объективен и обоснован.
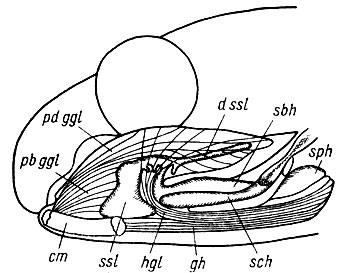
Рис. 16. Схема строения языка Rana temporaria. Графическая реконструкция с лимфатическими полостями, инъекцированными латексом Выбрасывание языка осуществляется совместным сокращением m. genioglosus и т. geniohyoideus. Pars basalis т. genioglossus (pb ggl), опираясь на нижнюю челюсть (cm), растягивает sinus sublingualis (ssl), в который поступает дополнительная порция лимфатической жидкости из sinus basihyoideus (sbh), который, в свою очередь, через sinus раrahyoideus (sph) и sinus ceratohyoideus (sch) связан с остальной лимфатической системой организма. Одновременно pars dorsalis m. genioglossus (pdggl) напрягает спинку языка и сжимает дистальную часть лимфатической полости, тогда как на ее дно давит сокращающийся m. geniohyoideus (gh). В результате давление в sinus sublingualis увеличивается, и она, распрямляясь, выпрямляет и выбрасывает язык. Сокращение вентрального поперечного слоя m. genioglossus проталкивает находящуюся в дивертикулах s. sublingualis лимфатическую жидкость в конец языка, что приводит к его удлинению. Остальные обозначения см. на рис. 23-28
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'