2.3.3. Функциональные изменения органов в ходе филогенеза
Принципы эволюционных преобразований органов в связи с их функцией разрабатываются с 1875 г., когда А. Дорн (Дорн, 1937) опубликовал основополагающую работу о смене функций. С тех пор число разнообразных принципов постепенно увеличивалось, и A. H. Северцов (1939), собравший, по-видимому, исчерпывающую литературу по этому поводу, опубликовал 13 принципов, сгруппировав их в две группы: 1) анцестральная функция меняется у потомков количественно; 2) анцестральная функция меняется качественно. А. Н. Северцов, вероятно, понимал, что 13 принципов характеризуют весьма разнородные ситуации, но подчеркивал, что детализация принципов позволяет подробнее проследить пути преобразования органов в ходе филогенеза.
Однако разработка проблемы продолжалась, и в данной работе кроме вышеизложенных принципов мультифункциональности и множественного обеспечения функций собрано 19 принципов.
I. Принцип интенсификации функций (Северцов, 1939; Plate, 1924) относится и к активным и к пассивным функциям. Выражается в усилении, совершенствовании механизма осуществления функции. Примерами интенсификации функций могут служить переход от гладкой к поперечно-полосатой мускулатуре, увеличение дыхательной поверхности легких в ряду наземных позвоночных, усиление защитных функций, таких как развитие покровительственной окраски, развитие вторичного панциря морских черепах и т. д. Подразумевается, что при интенсификации функций усиливается главная функция органа, которая качественно остается без изменений. Поэтому можно утверждать, что совершенствуется именно механизм (морфологические структуры), обеспечивающий выполнение этой функции. Так, у Pipidae в связи с вторично - водным образом жизни в бедных кислородом водоемах интенсифицируется функция легочного дыхания. В связи с этим легкие приобретают сложную ячеистую структуру, напоминающую альвеолярную, - случай уникальный среди Anura (de Jongh, 1972), и подъязычный аппарат увеличивается (см. рис. 19), обеспечивая вентиляцию сложных легких. Интенсификация функций, как подчеркивал А. Н. Северцов (1939), один из наиболее распространенных в природе и наиболее важных принципов изменения морфофункциональных систем, отражающих их прогрессивную эволюцию. Интенсификация функций часто, но не всегда связана со снижением мультифункциональности: подъязычный аппарат Pipidae обеспечивает усиление вентиляции легких, но утрачивает функцию транспортировки пищи. Он не способен к движениям в горизонтальной плоскости.
При переходе от макроморфологического уровня рассмотрения функциональной эволюции к более глубоким уровням (тканевому, клеточному, молекулярному) принцип интенсификации функций и связанная с ним специализация, выражающиеся в снижении мультифункциональности, выступают все более отчетливо. Вероятно, это обусловлено тем, что в гомеостатичной внутренней среде организма, когда все функции любой структурной единицы достаточно жестко фиксированы, дальнейшая адаптация этой системы определяется в первую очередь совершенствованием осуществления уже существующих функций, т. е. их интенсификацией.
Впервые на интенсификацию функций на тканевом уровне обратил внимание А. А. Заварзин (1986), выразивший это. правило в законе параллельных рядов тканевой эволюции именно тем, что в обеих ветвях животного царства - у первичноротых и у вторичноротых - совершенствование тканей внутренней среды и нервной ткани идет параллельно, но не дивергентно. Дивергентная эволюция особенно хорошо прослеживается на эпителиях (Хлопин, 1946), что связано с более разнообразным характером их связей с окружающей средой. Данные об эволюции органелл клеток еще слишком не полны, чтобы проследить па этом уровне соответствие или не соответствие их эволюции закону Заварзина.
На молекулярном уровне эта закономерность выступает еще более отчетливо. Наиболее важное обобщение по функциональной эволюции на молекулярном и биохимическом уровнях организации, которые в данном случае трудно разделимы, получено А. М. Уголевым (1982а, б; 1983а, б; 1985) в его теории функциональных блоков. В качестве таковых, согласно этой теории, выступают специализированные ферменты, например щелочная фосфатаза, треглаза, Na+ - К+-атефаза. Ферменты клеток про- и эукариот функционируют, естественно, во взаимодействии с вполне определенными рецепторами, почему и представляют собой "блоки". К подобным блокам, по-видимому, следует отнести и специализированные ферменты, управляющие транскрипцией, такие как ДНК-полимераза, PHK-праймаза, ДНК-геликаза, ДНК-топоизомеразы I и II и т. д. К функциональным блокам А. М. Уголев отнес и цикл трикарбоновых кислот Кребса, и гликоксилатный цикл. Блоки обеспечивают нормальное функционирование клеток и непосредственно или через функции клеток - более высоких уровней организации, включая целостный организм. Этим объясняется их универсальность, т. е. то, что они обнаруживаются в разных тканях и органах одного и того же организма и у разных организмов, подчас очень далеких друг от друга по своему таксономическому положению.
Так, гликоксилатный цикл, в ходе которого две молекулы ацетата превращаются в одну молекулу сукцината, обнаружен у бактерий, дрожжей, грибов, зеленых водорослей, высших растений, нематод и позвоночных. Na+ - К+-атефаза характерна, по-видимому, для всех поляризованных клеток организмов. То же относится к кислым протеазам, α-амилазе и другим пищеварительным ферментам. A. M. Уголевым (1985) собрано и обобщено огромное количество данных по распространенности и функционированию различных ферментных систем, относимых им к функциональным блокам. Широкое распространение блоков по различным таксонам указывает на то, что они формировались на самых ранних этапах эволюции жизни. Можно предполагать, что переход к многоклеточности был одним из путей поддержания эволюционной пластичности организма, давшим возможность более широкой комбинаторики функциональных блоков.
По-видимому, справедливы обе указанные цитируемым автором причины чрезвычайной эволюционной стабильности функциональных блоков: их монофункциональность, совершенно не свойственная структурам органного уровня, их коадаптированность, жесткая стабилизированность функциональных связей внутри блоков. Последнее - следствие эволюции динамических координаций - будет рассмотрено ниже. Moнo-, редко олигофункциональность - несомненное следствие интенсификации функций в гомеостатической внутриклеточной среде многоклеточного организма.
Высокая специализированность таких ферментов, как Na+ - К+-атефаза или эндонуклеаза I и т. п., обеспечивает надежность функционирования клетки как целого. Вместе с тем она, естественно, снижает эволюционную пластичность на молекулярном уровне организации. Сохранение этой пластичности может достигаться двумя основными путями - перекомбинацией блоков и изменением условий их функционирования, что может приводить к смене функций и (или) к дестабилизации блоков, обеспечивающей их дальнейшую эволюцию под действием движущего отбора по любому из принципов и типов филогенетических изменений.
Необходимо, однако, отметить, что моно- или олигофункционалыюсть специализированных ферментов на внутриклеточном уровне может оборачиваться мультифункциональностью на тканевом и органном уровнях. Так, опиоидные пептиды, связываясь с опиатными рецепторами ЦНС, не только подавляют боль, но и меняют психическое состояние, вызывая эйфорию. Вазопрессин, имея единственную функцию - стимуляцию аденилатциклазы, через посредство ц-АМФ на уровне целостного организма повышает кровяное давление, регулирует водный баланс и стимулирует долговременную память. Типичный блок в понимании А. М. Уголева (1985) - соматостатин, синтезирующийся в секреторных клетках кишечника, поджелудочной железе, спинном мозге, - не только ингибирует гормон роста, но также и продуцирование ряда других гормонов: тиреотропина, инсулина, глюкогона, гастрина, секретина. Вероятно, множественность действия гормонов обусловлена не только локализацией в организме их рецепторов, но и разнообразием этих рецепторов. Так, брадикинин имеет не только гипотензивное действие, осуществляющееся посредством усиления проницаемости стенок капилляров, но и вызывает сокращение гладкой мускулатуры, причем на мышцах кишечника это его действие сказывается сильнее, чем на мышцах матки. Не ясно, является ли физиологическое действие брадикинина на ЦНС как медиатора боли каким-либо образом связанным с его вазопрессорным эффектом или обусловлено иными механизмами действия этого регулятора.
Число подобных примеров может быть увеличено. Однако и приведенных достаточно, чтобы высказать предположение: отбор по опосредованным эффектам, точнее по интегральным функциям гормонов на более высоких уровнях организации, может служить эволюционным механизмом комбинирования и перестройки молекулярных блоков.
Имеются данные, позволяющие, с одной стороны, экстраполировать концентрацию функциональных блоков на более высокие уровни организации, прежде всего на органный, а с другой - дающие возможность понять механизм формирования подобных блоков. Т. Б. Голубева (1987) при исследовании эволюции онтогенеза слуха птиц показала, что развитие улитки и слухового эпителия, в частности его интенсивное разрастание и дифференциация волосковых клеток, стимулируется не только звуковыми сигналами наседки и других эмбрионов той же кладки, но и собственной вокализацией данного эмбриона, возникающей у выводковых птиц на пренатальных стадиях развития, а у птенцовых после вылупления. Иными словами, регуляция темпов развития внешними по отношению к организму факторами - сигналами других особей,, замещается авторегуляцией - собственными сигналами. Возникает обратная связь, консолидирующая данную функциональную систему и превращающая ее в блок, в понимании А. М. Уголева. Возникновение именно единого стабильного блока доказывается тем, что в эволюции птиц темпы индивидуального развития обеих систем - вокализации и интенсивного формирования слухового анализатора - меняются совместно, как единое целое. Так, у птенцовых развитие слуха приурочено к прозреванию птенцов, а у выводковых - к прорыву оболочки воздушной камеры яйца.
Наибольший интерес молекулярных биологов в последние десятилетия привлекает эволюция ДНК. Однако эта проблема очень сложна. В принципе, адаптивный смысл преобразований структуры ДНК, и в первую очередь последовательности нуклеотидов" должен быть установлен через исследования функций генов на фенотипическом уровне, начиная с транскрипции и кончая макроморфологией, физиологией и онтогенезом. Выяснить эти связи очень трудно, а там, где такая связь прослеживается, ее трудно увязать со всей структурой ДНК. Последовательность нуклеотидов должна отражать все особенности организации особей, в том числе рецессивные гены; гены, функционирующие на тех или иных этапах онтогенеза, т. е. и гены, работающие постоянно, и гены, активные лишь в определенные периоды развития; гены "домашнего хозяйства", "гены роскоши" и регуляторные гены; какие-то особенности генома, связанные с флуктуирующей изменчивостью; гены и их комплексы, связанные с видовой принадлежностью, включая гены, ответственные за сбалансированный полиморфизм как на белковом, так и на более высоких уровнях его проявления; изменения последовательности нуклеотидов, определяющие индивидуальную изменчивость, т. е. мутации и их комбинации; особенности структуры ДНК как фенотипического признака и тем самым объекта отбора и стохастических процессов; особенности структуры ДНК, сформировавшиеся в ходе предшествующей эволюции. Этот далеко не полный перечень приведен здесь лишь с целью напомнить о трудностях, стоящих на пути изучения эволюции ДНК. В подтверждение реальности этих трудностей достаточно напомнить о дискуссиях по поводу "эгоистической" ДНК и "молчащей" ДНК.
Пожалуй, не менее бурную дискуссию вызвали и представления нейтралистов, согласно которым эволюция ДНК (и белков) идет посредством накопления селективно нейтральных мутаций (Кимура, 1985; Ратнер, 1985; Ратнер и др., 1985; Ward, Skibinski, 1985 и др.). В настоящее время большинство исследователей признают существование нейтральных замен аминокислот в белковых молекулах, а следовательно, и нейтральность вызывающих эти замены мутаций.
Наиболее интересным представляется то, что филогенетические схемы, построенные на такой весьма ограниченной базе, как отдельные представители крупных таксонов (отрядов, классов, редко семейств), часто дают вполне удовлетворительное совпадение с филогенетическими схемами, построенными на базе макроморфологии традиционными методами (Рэфф, Кофмэн, 1986; Ayala, 1986).
Если схемы, построенные на основе анализа замен аминокислот, действительно объективны, т. е. не корректированы по традиционным схемам, то одного этого совпадения достаточно для признания селективной нейтральности замен. В том случае, если бы они хоть в какой-то степени отражали перечисленные выше селективно-значимые структурные особенности ДНК, подобное соответствие было бы невозможно. Выявилось бы именно своеобразие молекулярной эволюции (аналогично ее своеобразию на тканевом уровне). Только если замены действительно селективно нейтральны и не затрагивают адаптивных свойств и белков и ДНК, они могут служить интегральным отражением адаптивной эволюции всех уровней организации: от органного до молекулярного. Адаптивная эволюция всех этих уровней ставит пределы накоплению нейтральных мутаций и кодируемых ими замен аминокислот и тем самым формирует таксономические дистанции, адекватно отражающие своеобразие организации сравниваемых таксонов.
К сказанному необходимо добавить, что существование нейтральных мутаций, не противоречит представлению об отборе как о движущем факторе эволюции. Действительно, накопление нейтральных замен аминокислот в конце концов приведет к селективно-значимым изменениям структуры белка. С этого момента данный белок подвергается отбору, причем отбор, как и в любом случае отбора по фенотипической изменчивости, будет либо отсекать, либо подхватывать подобные изменения в зависимости от их адаптивной ценности. Аналогичное представление о селекционном механизме эволюции биологических макромолекул высказано А. М. Уголевым (1985). Можно даже предположить, что более мягкое и более плавное изменение структуры белков через накопление первично нейтральных замен благоприятнее с точки зрения отбора, чем мутационные перестройки, скачкообразно меняющие биологические свойства молекулы в результате изменений ее активного центра. Представление о "молекулярных часах", т. е. о равномерности во времени накопления нейтральных мутаций данного белка, не противоречит представлению о неравномерности темпов молекулярной эволюции (Kelly, Halladay, 1987; Li et al., 1987 и др.). Действительно, даже уже имеющиеся немногочисленные данные свидетельствуют о том, что каждый тип белков характеризуется собственным темпом накопления замен. Можно ожидать, что по мере увеличения количества исследованных белков картина будет усложняться.
С другой стороны, как сказано в разделе 1.2, сама нейтральность подобных мутаций может быть не первичной, а вторичной, т. е. представлять собой результат отбора по другим генам, компенсирующего первичную вредность подобных мутаций. А. Л. Мазин (1980) высказал предположение, что накопление селективно нейтральных (по отношению к последовательности аминокислот в белках) мутаций может быть селективно значимым по отношению к механизмам репликации и транскрипции ДНК. Наконец, А. Г. Креславский (устн. сообщ.) высказал предположение, что селективно-нейтральные мутации на самом деле могут иметь важное значение как ограничители возможности селективной перестройки тех белков, в которых они проявляются, или других белков, функционально связанных с мутантными. Таким образом, вопрос о реальном значении нейтральной эволюции остается открытым. С одной стороны, представления нейтралистов не могут быть приняты до тех пор, пока не будет доказана селективная нейтральность мутаций в момент их возникновения. С другой - нейтральность хотя бы части вновь возникающих мутаций не может быть опровергнута до тех пор, пока не доказаны все вышеизложенные гипотезы.
II. Принцип фиксации фаз (Северцов, 1939) описывает частный случай интенсификации функций, когда усиливается лишь одна из ряда фаз сложного движения органа. Например, при интенсификации бега - переход от стопохождения к пальцехождению. Действительно, толчок пальцами о грунт - последняя фаза опоры конечности при локомоции стопоходящих животных. При беге этих животных она становится единственной. Интенсификация функции пальцев в этой фазе движения и приводит к пальцехождению. Фиксация фаз, как и интенсификация функций, ведет к снижению мультифункциональности: специализированные конечности пальцеходящих млекопитающих, особенно копытных, не пригодны для схватывания, лазания и т. п.
III. Принцип выпадения промежуточных функций, как отмечал A. H. Северцов (1939), малораспространенный. Как и предыдущий, он связан с интенсификацией функций. Этот принцип выражается в упрощении системы, обеспечивающей функцию, в совершенствовании аппарата, осуществляющего ее. Так, в ряду позвоночных квадратная кость утрачивает функцию элемента, связывающего нижнюю челюсть с черепом, в результате чего сочленение стало более прочным и функция челюстей интенсифицировалась.
IV. Принцип субституции органов (Kleinenberg, 1886, цит. по Северцову, 1939). Согласно этому принципу в процессе филогенеза (и онтогенеза, по Клейненбергу) орган, выполнявший в организме определенную функцию, исчезает, но ту же функцию начинает выполнять другой по происхождению, т. е. аналогичный орган, который может находиться в ином месте. Общеизвестны примеры субституции хорды позвоночником, зубов - роговым клювом (черепахи, птицы), листовых пластинок - филлодиями у Acacia русnantha и т. д.
Нетрудно заметить, что субституция органов основана на принципе множественного обеспечения биологически важных функций. Действительно, у рептилий в захватывании пищи участвуют и зубы и роговые чешуи губного ряда, в фотосинтезе участвуют и листовые пластинки и черешки листьев. Редукция одного из компонентов системы, обеспечивающей функцию, сопряжена с интенсификацией функций другого компонента, причем, как почти всегда происходит при интенсификации функций, мультифункциональность субституирующего органа оказывается ниже, чем субституируемого.
V. Принцип физиологической субституции (Федотов, 1925, цит. по Северцову, 1939) отличается от предыдущего тем, что субституирующий орган функционально замещает субституируемый, уподобляясь ему морфологически, но субституируемый орган при этом не исчезает. Примером физиологической субституции, согласно Д. М. Федотову, может служит превращение у офиур целома в выводковую камеру. Так, у Segophiura nodosa выводковыми полостями служат бурсы - 5 пар полостей, открывающихся во внешнюю среду. Кроме функции выводковой камеры бурсы выполняют функции дыхания и половую. У Ophionotus hexactis личинки вынашиваются не в бурсах, а в целомической полости, тогда как остальные функции бурс, как и строение этих органов, остаются неизменными. Нетрудно заметить, что в данном случае описан частный случай субституции функций (см. ниже). Показательно, что при этом мультифункциональность бурс снижается, а мультифункциональность целомической полости возрастает, хотя общее число функций обеих полостей остается неизменным.
VI. Принцип уменьшения числа функций (Северцов, 1939). Как подчеркивалось выше, при интенсификации главной функции органа мультифункциональность его снижается. A. H. Северцов (1939) возвел это положение в ранг специального принципа, согласно которому "...при филогенетическом усилении главной функции какого-либо органа общее число функций этого органа у потомков уменьшается" (с. 380). Цитируемый автор подчеркнул, что уменьшение числа функций приводит к снижению эволюционной пластичности органов, т. е. к их специализации.
Вышеизложенными шестью принципами A. H. Северцов (1939) ограничил число принципов, в которых, согласно его классификации, функция меняется количественно.
VII. Принцип расширения функций (Plate, 1912). Л. Плате (1912) обратил внимание на то, что в процессе прогрессивного развития органа он может достигать такой степени дифференциации, при которой берет на себя дополнительные (второстепенные) функции, ранее ему не свойственные. Как пример расширения функций Л. Плате приводил приобретение жаберной полостью пластинчатожаберных моллюсков функции выводковой камеры. По-видимому, использование офиурами целома в качестве выводковой камеры также можно рассматривать как расширение функций. A. H. Северцов показал, что эволюция парных плавников у кистеперых рыб шла по принципу расширения функции: их дополнительная функция - опора на субстрат. Так же и грудные плавники летучих рыб приобрели дополнительную функцию полета. Принцип расширения функций, представляющий собой антитезу принципу уменьшения функций, обусловливает возрастание мультифункциональности органов. Качественный характер изменения функций выражается именно в приобретении дополнительных новых функций.
VIII. Принцип смены функций (Дорн, 1875, на русск. яз., 1937). Согласно А. Дорну, ослабление главной или усиление второстепенной функции приводит к такой перестройке морфологии органа, при которой одна из его второстепенных функций становится главной, а главная функция у предков либо становится второстепенной, либо исчезает. Примеры смены функций - одного из наиболее распространенных принципов функциональной эволюции органов очень многочисленны: превращения ходильных конечностей в ласты у вторичноводных млекопитающих, третьей жаберной дуги в челюсти, яйцеклада в жало у рабочих пчел и т. п. Утрата или ослабление основной функции предков, несомненно, приводит к качественной перестройке набора функций органов. В то же время нетрудно заметить связь смены с интенсификацией функций. Превращение одной из второстепенных функций в главную неизбежно происходит через интенсификацию этой функции.
IX. Принцип субституции функций (Северцов, 1939). "Функция органа предков, эволюционирующего согласно этому типу филогенетического развития, замещается (субституируется) у потомков другой функцией, биологически ей равноценной, но выполняемой другим органом, расположенным в другой части тела животного и развившимся из другого зародышевого зачатка". Так, при переходе от локомоции при помощи парных конечностей (у ящериц) к ползанию (у змей) позвоночник и ребра с их мускулатурой функционально замещают редуцирующиеся конечности. У ластоногих и китообразных функция термоизоляции тела, выполнявшаяся шерстным покровом, замещается функцией термоизоляции, выполняемой жировым слоем подкожной клетчатки. Последний пример, приведенный A. H. Северцовым (1939, с. 403), еще в большей степени, чем предыдущий показывает, что субституция органов, физиологическая субституция и субституция функций связаны между собой практически нечувствительными переходами. Не удается показать, почему замещение теплоизолирующей функции шерсти той же функцией жирового слоя представляет собой именно субституцию функций, а не органов. К тому же у тюленей, у которых шерстный покров сохраняется, но его терморегуляторное значение очень мало, а жировой слой именно в связи с термоизоляцией развит очень сильно, видимо, можно говорить о физиологической субституции.
Относительно всех трех принципов можно говорить как о принципах, основанных на множественном обеспечении функций и естественно связанных между собой.
X. Принцип активации функций (Северцов, 1939) описывает, по-видимому, довольно редкие в природе случаи превращения пассивной функции в активную. Таков переход от неподвижного прикрепления челюстей у ящериц к их независимой от нейрокрана подвижности, активному кинетизму черепа у змей, обусловленному первичным для Tetrapoda кинетизмом черепа (Иорданский, 1985). Таково же, вероятно., преобразование quadratum в columella auris. В данном случае, как и во многих других принципах, прослеживается тесная связь с принципом интенсификации функций.
XI. Принцип иммобилизации функций (Северцов, 1939) представляет собой альтернативу принципу активации. Он констатирует, что активная функция в ряде случаев может становиться пассивной. Так, у акуловых рыб palatoquadratum верхняя челюсть подвижна относительно осевого черепа - гиостилия, у химеровых рыб верхняя челюсть срастается с осевым черепом и теряет подвижность - аутостилия. Следует, однако, отметить, что иммобилизация функций отнюдь не связана с редукцией органа или ослаблением функции. Напротив, этот принцип также связан с интенсификацией: прирастание верхней челюсти к осевому черепу у химер привело к укреплению челюстей и к их усилению. Химеры могут раздавливать раковины моллюсков, панцири ракообразных и т. п. Обращает внимание, что описанный здесь по А. Н. Северцову (1939) пример иммобилизации функций очень близок к фиксации фаз. Действительно, среди довольно сложных движений челюстей акуловых, служащих для разрезания добычи, при которых верхняя и нижняя челюсти смещаются друг относительно друга, а пилообразные зубы разрезают ткани жертвы подобно ножам косилки, есть фаза схватывания добычи, когда зубы внедряются в тело жертвы. Эта фаза и усиливалась, оставшись единственной у химер.
XII. Принцип симиляции функций (Северцов, 1939). Как считал A. H. Северцов (1939), симиляция - уподобление функций - (может быть, правильнее сказать органов и функций) - весьма распространенный тип филогенетических преобразований органов. В качестве примеров симиляции он приводил уподобление формы, а также функции шейных и поясничных позвонков и ребер туловищным при возникновении змей от ящерицеобразных предков. Действительно, удлинение тела и субституция функции бега ползанием привели к удлинению тела, увеличению морфологического сходства между позвонками и удлинению шейных и поясничных ребер до размеров грудных. Нетрудно заметить связь и этого принципа с интенсификацией функций, однако не тех позвонков и ребер, функции и строение которых становятся все более подобными друг другу, а посткраниального осевого скелета в целом, все элементы которого совокупно обусловливают локомоцию змей.
XIII. Принцип разделения органов и функций (Северцов, 1939). Примерами этого типа преобразований органов могут служить разделение боковых складок тела предков челюстноротых на грудные и брюшные плавники; разделения непарной плавниковой складки на спинные, хвостовой и анальный плавники; многоэтапная дифференциация пищеварительного тракта на отделы и т. п. Следует отметить, что в применении к целостному организму этот принцип был открыт Мильн Эдвардсом, согласно которому в процессе прогрессивной эволюции организмы дифференцируются. Впрочем, и его нельзя считать первооткрывателем. К. Бэр (1828, цит. по Дарвину, 1939) предлагал оценивать уровень организации по степени физиологического разделения труда между частями организма. С другой стороны, принцип разделения органов и функций очень близок к принципу выпадения промежуточных функций. Оба эти принципа описывают дифференциацию систем.
Вышесказанным исчерпаны принципы и типы филогенетических изменений органов, собранные A. H. Северцовым. В изданной в 1954 г. монографии В. А. Догель подытожил свои многолетние исследования проблемы филогенетической дифференциации организма, в нее вошли три разработанных им принципа филогенетических изменений органов.
XIV. Принцип полимеризации. Согласно этому принципу в филогенезе часто наблюдается увеличение числа гомодинамных и гомономных образований, связанное с интенсификацией функции системы, подвергавшейся полимеризации. Примерами полимеризации могут служить, например, вторичное разделение жаберных щелей у ланцетника, доводящее их число иногда до 90 пар и более, удлинение позвоночника змей, вторичная сегмнтация тела у пиявок и т. п. Во всех подобных случаях четко прослеживается интенсификация функций - суммарная дыхательная поверхность 90 пар щелей, больше, чем 7 пар у предполагаемых предков ланцетника; подвижность позвоночника змей больше, чем у ящериц, и т.п. Вместе с тем полимеризация, по-видимому, не приводит сама по себе к снижению мультифункциональности, так как не ведет к перестройке каждого элемента полимерной системы. С другой стороны, полимеризацию можно рассматривать как один из путей возникновения систем множественного обеспечения функций. Действительно, компоненты полимерной системы морфологически автономны и функционально между собой не связаны, напротив, каждый из них вносит свой вклад в обеспечение общей для них функции.
Видимо, следует различать два случая возникновения полимерных систем: вышеприведенные примеры относятся к полимеризации систем, уже существовавших у предков. Однако можно привести ряд примеров возникновения полимерных признаков, которые являются следствием возникновения эволюционно нового, отсутствовавшего у предков признака. К таким первичным полимерным системам, вероятно, можно отнести глазки Гессе ланцетника, мантийные глазки у Pecten, щупики на "губах" у личинок бесхвостых амфибий. Ярким примером возникновения первичной полимерной системы может служить описанный ниже пример возникновения точечного рисунка на надкрыльях у жуков-листоедов (Креславский, 1977). В подобных случаях едва ли можно говорить об интенсификации функций. Ведь функция световооприятия ранее, т. е. у предков Pecten, не существовала, не существовали и структуры, ее обеспечивающие.
Таким образом, по-видимому, лишь вторичная полимеризация может рассматриваться в ряду других принципов и типов филогенетических изменений органов. Однако и в такой интерпретации этот принцип не укладывается в предложенную А. Н. Северцовым классификацию. Действительно, поскольку при полимеризации происходит интенсификация функций, этот тип следует отнести к принципам, описывающим количественные изменения функции.
XV. Принцип олигомеризации (Догель, 1954) характеризует тенденцию, альтернативную полимеризации, - уменьшение числа компонентов полимерных систем. Примерами олигомеризации могут служить: уменьшение числа жаберных дуг в ряду примитивных челюстноротых - от исходных 11 пар через 9 пар, свойственных Palaeonisci, до 7 пар у примитивных акул и 5 пар - высших акул; уменьшение числа лучей парных конечностей у кистеперых рыб по сравнению с лучеперыми и у Tetrapoda по сравнению с кистеперыми; уменьшение числа покровных костей черепа у высших рыб по сравнению с осетровыми или у рецентиых амфибий по сравнению со стегоцефалами.
Все приведенные примеры отражают вторичную перестройку полимерных систем, но пути этой перестройки различны. Олигомеризации может происходить за счет редукции частей полимерной системы, как это наблюдается в процессе эволюции парных конечностей и отчасти висцерального скелета; путем слияния элементов, как это часто наблюдается в покровных окостенениях черепа (Лебедкина, 1979); путем перестройки части элементов по принципу, например, смены функций, как это произошло с челюстной и гиоидной дугами, возникшими из жаберных. Таким образом, принцип олиогмеризации описывает лишь общую картину преобразования полимерных систем, но не конкретные пути их эволюции. Видимо, для объяснения этих путей В. А. Догелем (1954) введен принцип компенсации функций.
XVI. Принцип компенсации функций (Догель, 1954). При олигомеризации пути преобразования элементов, выходящих из состава полимерной системы, разнообразны, но остающиеся в системе элементы продолжают выполнять ту же функцию, что и у предков. Поскольку эволюция не может идти по пути снижения приспособленности, сохранившая свою функцию часть полимерной системы должна выполнить эту функцию не хуже, чем выполняла ее вся система у предков. Поэтому при олигомеризации сохраняющиеся части полимерной системы компенсируют утрату функции редуцированных или сменивших функцию частей. В этом, собственно, и заключается компенсация функций по Догелю, связанная, как это нетрудно заметить, с интенсификацией функций сохранившихся компонентов и тем самым со снижением мультифункциональности.
Истоки принципа компенсации функций можно, видимо, проследить вплоть до закона компенсации Э. Жоффруа Сент-Илера. Однако для понимания значения этого принципа важнее более широкая трактовка, приданная ему H. H. Воронцовым (1961, 1967). Компенсация может происходить не только при олигомеризации полимерной системы, но и между компонентами биологически координированных систем, совокупно обеспечивающих ту или иную функцию. Например, мускулистый желудок птиц компенсирует редукцию у них зубов; большая кожная вена амфибий, обеспечивая кожное дыхание, компенсирует слабость легочного газообмена. Развитие хлорофилла в стеблях кактусов компенсирует редукцию их листьев и т. д. В таком виде принцип компенсации в большей степени, чем у В. А. Догеля (1954), напоминает представления Э. Жоффруа Сент-Илера. Но в то же время он показывает одно из важных свойств организма как целого: снижение мультифункциональности - олигофункциональность, если так можно выразиться, ведет к снижению эволюционной пластичности признаков - специализации. Однако компенсация функций, благодаря которой утрата функций (и органов) может компенсироваться возникновением аналогичных функций других частей организма, как показал Н. Н. Воронцов (1961, 1967), ведет к переспециализации. Иными словами, организм в целом функцию не oутрачивает, сохраняя тем самым эволюционную пластичность.
XVII. Принцип смены адаптивных норм реагирования (Шмальгаузен, 1969) представляет собой расширение принципа смены функций до рамок целостного организма. При достаточно глубоких изменениях условий среды меняются, собственно, не отдельные признаки, а все соотношения организма со средой - норма его реагирования на внешние условия. Тем самым создается новая - основа для дальнейшей эволюции. Однако, возводя в ранг принципа представление о том, что в ходе филогенеза возникают новые адаптивные нормы реагирования, обсуждаемый принцип отличается от всех вышеизложенных тем, что не позволяет анализировать процесс становления этих новых норм даже на том описательном уровне, который свойствен всем другим изложенным здесь oпринципам и типам филогенетических изменений органов в связи с их функцией.
XVIII. Принцип эволюционной стабилизации функций (Кокshaysky, 1973). Согласно этому принципу при изменении условий функционирования, например при изменении формы крыла, одна из частей организма дополнительно меняется таким образом, чтобы сохранить функцию органа неизменной.
XIX. Принцип концентрации функций (Кокшайский, 1980) характеризует процесс топографической локализации функции в какой-либо части органа. Например, у скумбриеобразных рыб основное значение в локомоции имеют движения именно хвоста, а не задней части тела. В связи с этим, видимо, следует отметить, что из кратких описаний принципа (Кокшайский, 1980) не удается выявить разницу между этим принципом и принципом фиксации фаз, который также характеризует преимущественное развитие и интенсификацию функции той части органа, которая выполняет важнейшую фазу движения.
Подводя итоги вышеизложенным принципам перестройки морфофункциональных систем в ходе филогенеза, необходимо прежде всего обратить внимание на то, что среди этих 19 принципов нет ни одного, описывающего процессы редукции. Редукцтия органов вообще довольно слабо изучена, но отсутствие функциональных подходов к редукции свидетельствует скорее об односторонности подхода, чем о несущественности принципов редукции для понимания филогенеза. Лишь А. Н. Северцов (1939) отметил, что редукция органов может происходить при утрате органом его биологического значения в силу изменения условий среды, при субституции органов, иногда при смене функций и уменьшении их числа.
Принципы, описывающие прогрессивное развитие органов в связи с их функцией, крайне разнородны как по частоте встречаемости, так и по значению. Например, практически во всех типах прогрессивного изменения органов в том или ином виде присутствует интенсификация функций, тогда как эволюционная стабилизация функций или выпадение промежуточных функций, видимо, достаточно редки. При изложении принципов полимеризации и олигомеризации было подчеркнуто, что эти принципы сами по себе пе элементарны: под одним и тем же названием подразумеваются разные по происхождению и эволюционному значению процессы.
Классификация принципов согласно предложенному А. Н. Северцовым критерию различения количественных и качественных изменений функций (Северцов, 1939; Тимофеев-Ресовский и др., 1969) трудно приложима к таким принципам, как полимеризация и олигомеризация. Кроме того, ряд принципов, очень сходных между собой (например, субституция органов и субституция функций), оказываются в разных разделах. Наконец, можно ожидать, что будут формулироваться новые принципы (Шкробатов, 1986) и их система будет становиться все более запутанной. Иными словами, они представляют собой не принципы, а более или менее часто встречающиеся ситуации и различные аспекты рассмотрения одних и тех же ситуаций. Поэтому представляется целесообразным подойти к уже описанным принципам не с позиций их классификации, а с позиций анализа их взаимосвязей и путей становления.
По-видимому, можно различать три типа морфологических систем, преобразования которых описывают вышеизложенные принципы: 1) единая морфологическая структура (органелла, клетка, орган, организм). В основе возможности преобразования таких систем лежит принцип мультифункциональности (I тип морфофункциональных систем); 2) набор функциональных систем, совокупно обеспечивающих выполнение той или иной общей для них функции, но не связанных между собой непосредственными функциональными связями. В основе преобразования таких систем лежит принцип множественного обеспечения функции (II тип морфофункциональпых систем); 3) промежуточное положение занимают более или менее сложные морфофункциопальные системы, части которых совокупно обеспечивают выполнение той или иной функции, но в то же время непосредственно связанные между собой функционально и часто морфогенетически (III тип морфофункциональных систем). Примером может служить пищеварительная система (Уголев, 1985).
Для преобразования мультифункционалыюй системы мыслимы три варианта: отбор идет по главной функции, отбор идет по второстепенной функции и отбор идет и по главной, и по второстепенной функциям. При отборе по главной функции происходит ее интенсификация, сопровождаемая уменьшением числа функций и специализацией органа. При отборе по второстепенной функции происходит ее интенсификация и через фазу существования двух главных функций - смена функций. Чем слабее отбор по анцестральной главной функции или чем интенсивнее отбор по второстепенной, тем быстрее происходит смена функций. При одновременной интенсификации и главной и второстепенной функций в системах промежуточного типа происходят выпадения промежуточных функций, или разделения органов и функций, или при отсутствии возможности осуществления какого-либо из этих принципов (высокой морфофункционалыюй интеграции системы) возникает ситуация существования двух главных функций.
Более интересен случай, когда отбор, продолжаясь по главной функции, меняет свое направление. В подобных случаях возникает фиксация фаз, концентрация, стабилизация и иммобилизация функций или их активация. По-видимому, перечисленные принципы не исчерпывают всех ситуаций, которые возникают при изменении направления отбора по главной функции. Особое место занимает вторичная полимеризация. Она возникает в тех случаях, когда отбор по главной функции действует на уже более или менее полимерную систему гомодинамных или гомономных органов. Полимеризация сопровождается симиляцией функций и приводит, по сути дела, к возникновению системы множественного обеспечения функций.
Если структура полимерной системы не поддерживается специфическим вектором отбора, стабилизирующим ее, то дальнейшее преобразование подобной системы (ее олигомеризации), видимо, неизбежно. Действительно, в силу протяженности полимерной системы в пространстве ее компоненты оказываются в разных условиях функционирования. Например, поток воды через передние и задние жаберные щели проходит с разной скоростью (Северцов, 1980а, б); сколь бы ни были похожи друг на друга позвонки змей, туловищные позвонки и позвонки задней трети тела играют разную роль в локомоции животного. На элементы системы, оказавшиеся в разных условиях функционирования, отбор действует в разных направлениях, в результате чего система дифференцируется. Типы (пути) филогенетических изменений тех элементов полимерной системы, которые выходят из ее состава, видимо, могут быть любыми, за исключением восстановления их анцестральной главной функции, но для элементов, сохраняющих эту функцию, ее интенсификация неизбежна. Она и обеспечивает компенсацию функций преобразующихся частей системы.
Компенсация функций, по Н. Н. Воронцову (1961, 1967), может происходить, видимо, только в морфофункциональных системах промежуточного типа, в которых множественное выполнение той или иной функции обеспечивается или дополняется функциональными связями между частями системы. Следует отметить, что принципы функциональной эволюции подобных систем, динамических координаций по терминологии А. Н. Северцова (1939) (см. ниже), наименее изучены, но, вероятно, наиболее интересны.
Филогенетическая редукция морфологических структур происходит, по-видимому, в тех случаях, когда их главная функция утрачивает свое значение и отбор по ней ослабевает или прекращается. Примерами подобной редукции может служить рудиментация конечностей у безногих ящериц (Северцов, 1939) или редукция hypobranchiale III у личинок хвостатых амфибий (Северцов, 1974). Срастание концов, поддерживающих жаберные дуги ceratobranchialia, и редукция окостенений жаберных дуг обусловили то, что при движении опускания дна ротовой полости большое значение приобрела эластичность связи hypobranchiale с copula (рис. 20), hypobranchiale III снижало подвижность жаберных дуг (ceratobranchialia) относительно копулы. В результате связь между hypobranchiale III и копулой редуцировалась. Но после утери связи hypobranchiale III утратило функциональное значение, оно не участвовало в укреплении жаберных дуг и не препятствовало их движениям. В результате редукция этого элемента шла крайне медленно. У Lysorophidae, Lepospondyli этот элемент еще сохранялся (Romer, 1933а, б), причем был сочленен с копулой (Северцов, 1974). Следовательно, он начал редуцироваться у предков Urodela не позднее юры. У современных примитивных Urodela рудимент этого хряща присутствует постоянно, например у Rotiodon sibiricus, часто встречается у Hynobius keyserlingii, а у Salamandridae он встречается в качестве очень редкой вариации.
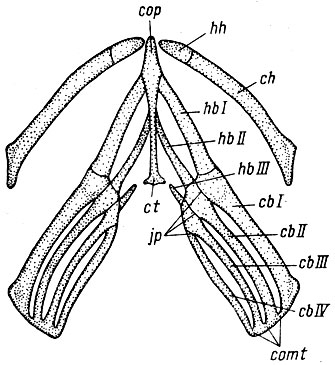
Рис. 20. Подъязычно-жаберный скелет личинки Ranodon sibiricus: cb I, II, III, IV - ceratobranchialia I, II, III, IV, ch - ceratohyale, ct - cartilago triangularis, comt - comissurae terminales, cop - copula, hb - I, II, III - hypobranchialia I, II, III (hb III рудиментарно), hh - hypohyale, jp - juncturae proximales
Следует, однако, отметить, что рудиментация, связанная, как показано выше, с прекращением отбора по главной функции, по-видимому, не препятствует осуществлению редуцирующимися структурами их второстепенных функций. Напротив, можно предполагать, что длительное существование рудиментов не только в онтогенезе, но и у взрослого организма, обусловлено именно их ролью как носителей второстепенных функций, в той или иной мере поддерживаемых отбором. При отборе против признака, возникающем, когда главная его функция не только утрачивает адаптивное значение, но и становится вредной, происходит афанизия (Северцов, 1939) - полное исчезновение органа.
Таким образом, с функциональной точки зрения могут существовать три типа редукции: 1) орган, утратив главную функцию, сохраняет второстепенные и поэтому сохраняется в виде рудимента, например аппендикс человека или гальтеры мух; 2) утратив главную функцию, орган теряет и второстепенные. В подобных случаях орган подвергается более быстрой и полной редукции, сохраняясь в онтогенезе, видимо, лишь в качестве элемента формообразовательного аппарата, а у взрослого организма - в качестве индивидуальной вариации, тем более редкой, чем дальше зашла его редукция. Примерами могут служить: редукция hypobranchiale III хвостатых амфибий, редукция конечностей у змей и безногих ящериц и т. п.; 3) наконец, при афанизии, когда признак вреден для функционирования включающей его системы, орган может сохраняться лишь как элемент формообразовательных зависимостей, т. е. только как возникающая и исчезающая в ходе онтогенеза закладка, по наличию которой мы и судим о том, что этот орган существовал у предков данной систематической группы. Наиболее известный пример - закладка жаберных щелей у Amniota.
Преобразование систем множественного обеспечения биологически важных функций определяется той же ситуацией, что и преобразование полимерных систем - неодинаковой ролью компонентов в обеспечении общей для них функции. Так, у бесхвостых амфибий легкие могут обеспечивать от 35 (Rana temporaria) до 100% (Bombina bombina) потребления кислорода и от 12 (Rana temporaria) до 53% (Pelobates fuscus) выделения углекислого газа. У хвостатых амфибий легкие могут обеспечивать от 47 (Hynobius keyserlingii) до 0% (Onychodactilus fischeri - безлегочная форма) потребления кислорода, но практически не участвуют в выделении углекислого газа. У Onychodactilus fischeri через кожу может поступать 100% необходимого организму кислорода. В выделении углекислоты кожа бесхвостых амфибий имеет ведущее значение, обеспечивая от 50 (Bombina bombina) до 100% (Bufo bufo) этой функции. У всех исследованных видов Urodela вся углекислота (100%) может выводиться через кожу (Северцов, Соколов, 1972, 1975, 1976).
У хвостатых амфибий кожа из-за несовершенства механизма вентиляции легких играет большую, чем у бесхвостых, роль в процессе газообмена. Поэтому в благоприятных условиях среды, при высокой влажности воздуха, происходит интенсификация дыхательной функции кожи, а легкие, утрачивая свое функциональное значение, редуцируются, как это произошло у Mertansiella саuсаsica Onychodactilus fischeri, у всех Plethodontidae и ряда других видов. Пользуясь терминологией принципов и типов филогенетических изменений органов, можно сказать, что у всех этих форм произошла субституция функций - кожа берет на себя функции легких.
У гребенчатого тритона (Triturus cristatus) через слизистую ротоглоточной полости может поступать весь необходимый животному кислород, у обыкновенной саламандры (Salamandra salamandra), относящейся к другому роду того же семейства, через слизистую ротоглоточной полости поступает не более 15% кислорода, примерно 15% поступает через легкие и почти 70% - через кожу. Можно говорить, что в данном случае происходит физиологическая субституция - морфология всех этих органов не меняется. На самом же деле мы сталкиваемся либо с интенсификацией дыхательной функции ротоглоточной полости (для тритона), либо с одним из этапов субституции легких кожей (для саламандры) (Возможность двоякой интерпретации обусловлена неоднозначностью прочтения сравнительного ряда). По-видимому, такова же основа и субституции органов. При интенсификации функций одного из них другой редуцируется. Следует отметить, что подобные процессы могут происходить и в системах промежуточного типа. Таким образом, все принципы и типы, связанные с редукцией одних органов и развитием других, основаны па множественном обеспечении функций.
Как следует из вышеизложенного, принципы и типы филогенетических изменений органов плавно переходят друг в друга или описывают различные стороны одного и того же процесса изменения данного органа. Поэтому правильнее говорить о более или менее распространенных типах функциональных преобразований структур, но не о принципах. Обращает на себя внимание, что все вышеописанные типы не способствуют повышению мультифункциональности морфологических систем. При интенсификации функции мультифункциональность обычно снижается, во всех остальных случаях, за исключением расширения функций (см. ниже), она не повышается, и лишь при полимеризации за счет формирования новых структур возникает база для дальнейших преобразований, т. е. пластичность организации повышается без увеличения мультифункциональности. Поэтому весьма важно рассмотреть механизмы повышения эволюционной пластичности организации. Они могут быть двоякими: либо возникает новая система, повышающая пластичность организации, так как она, обладая некоей мультифункциональностью и вступая во взаимодействия с другими системами, в состоянии меняться в разных направлениях, либо уже существующая система приобретает ранее не свойственные ей функции (расширение функций).
Первый случай описан А. Г. Креславским (1977) на примере возникновения рисунка на надкрыльях у жуков-листоедов. Неравномерность распределения пигмента (рисунок) возникает как побочный эффект эволюции структуры надкрыльев: пигмент скапливается в углублениях, возникающих при формировании внешнего слоя надкрыльев, образованного (в филогенезе) разросшимися и сросшимися жилками крыла. Первично такой рисунок, состоящий из мелких, не упорядоченных по расположению или не собранных в более или менее правильные ряды точек, не имеет собственного функционального (адаптивного) значения. Он является побочным результатом морфогенетических процессов и интенсивности метаболизма в функционирующих надкрыльях. Однако, раз возникнув, рисунок теряет свою безотносительность к внешней среде. Он может либо маскировать, либо демаскировать животное на окружающем фоне. Как только рисунок начинает оказывать влияние на приспособленность, он приобретает самостоятельное функциональное значение - собственную функцию, по которой уже может идти (и идет) отбор, приводящий к интенсификации данной функции: сигнальной (привлечение особей другого пола или предостерегающей окраски и т. п.) или покровительственной, т. е. маскирующей жука на окружающем фоне. При возникновении отбора по функции начинается эволюция рисунка: он становится упорядоченным и не связанным со структурой поверхности надкрыльев или исчезает, если становится вреден, или утрачивает функциональное значение.
Возникновение новых, не функциональных структур вследствие коррелятивных преобразований функциональных признаков ведет к появлению новых функций в том случае, если при данной экологической ситуации признак вступает в связь с внешней средой или, возможно, с другими признаками организма. Следует отметить, что, по сути, возникновение точечного рисунка на надкрыльях соответствует возникновению первично полимерной системы, которая затем подвергается олигомеризации, как и показал А. Г. Креславский (1977). Трудно сказать, во всех ли случаях первично полимерные системы имеют в своей эволюции дофункциональную фазу, т. е. возникают в результате коррелированных преобразований других систем, или часть из них - результат непосредственного действия отбора. Последний случай мыслим, если при отсутствии макроструктур, способных адаптивно реагировать на изменения внешней среды, отбору подвергаются структуры более глубоких уровней организации - тканевого, клеточного и т. п. Так, на основе данных В. Е. Соколова (1964) можно предполагать, что возникновение ороговений эктодермальных клеток кожи амфибий - следствие отбора на тканевом уровне: отмершие ороговевшие клетки перестали слущиваться и сформировали роговые бляшки - полимерная система.
Обнаруженное А. Г. Креславским (1977) явление решает один из важных вопросов теории эволюции: проблему возникновения новых признаков. По своей сути отбор может иметь дело лишь с существующими признаками, обладающими функциями. Иными словами, отбор на основе наследственной изменчивости может менять лишь уже существующие признаки. Поле его деятельности в этом отношении почти безгранично, но тем не менее существовала проблема, часто выдвигавшаяся противниками селекционизма: как могут возникать совершенно новые признаки, т. е. признаки, не являющиеся результатом преобразования признака предков под действием отбора? Представление о формировании новых признаков вследствие коррелятивных преобразований других структур решает этот вопрос в пользу дарвинизма и решает его окончательно.
Второй способ возникновения новых функций наблюдается, когда орган приобретает не свойственную ему ранее функцию путем разделения уже существующей на две или более самостоятельные функции без изменения структуры выполняющего их органа. Этот случай удобно рассмотреть на примере расширения функций, приведенном А. Плате (Plate, 1924). Жаберная полость двустворчатых моллюсков выполняет две основные функции: дыхания и транспорта пищи. Обе они зависят от движения ресничек ресничного эпителия жаберных мешков. Ресничный эпителий создает ток воды, который одновременно и омывает жабры, и несет органическую взвесь - пищу моллюска к ротовому отверстию. Но у ряда Lamellibranchiata жаберная полость стала также выводковой камерой: молодь проходит в ней весь период онтогенеза, от яйца до молодого животного.
Несколько более сложный пример аналогичного расширения функций как следствия возрастания функциональной нагрузки на признак, создающего некую избыточность функциональных возможностей органа, дает развитие механизма действия челюстей личинок Anura. Переход головастиков ,к фитофагии - питанию обрастаниями, стимулированный обитанием в горных потоках (Шмальгаузен, 1964), привел к вычленению из первично единой нижней челюстной дуги infrarostralia и формированию suprarostralia, а также механизма движений этих отделов челюстей. Первичным, по данным В. Б. Никитина (1980), был механизм именно скобления. Но это движение обусловило столь развитую подвижность губных хрящей, что возникло и движение кусания - вторая функция челюстного аппарата (рис. 21). В данном случае интересно то, что функция окобления сформировалась как следствие смены функций нерасчлененной челюсти того же типа, какой был свойствен личинкам Urodela (см. рис. 14, 16), т. е. вследствие отбора по второстепенной функции, а кусание - как результат дальнейшей интенсификации этой же, ставшей уже главной функции.
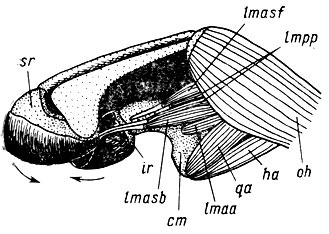
Рис. 21. Челюстной аппарат головастика Pelobates fuscus Стрелками указаны движения верхнего (suprarostrale - sr) и нижнего (infrarostrale - ir) губных хрящей, одетых роговыми чехлами. Движение верхнего губного хряща, связанного упругой соединительной тканью с трабекулами, осуществляется сокращением m. levator mandisulae anterior subexternus (lmasb) и одной из порций т. levator mandibulae posterior profundus (lmpp). Движения нижнего губного хряща обусловлены поворотами нижней челюсти - меккелевых хрящей (cartilago mecceli - cm), которыми управляют m. hyoangularis (ha) и m. quadratoangularis (qa), отводящие нижнюю челюсть и разводящие передние концы меккелевых хрящей, а так же m. levator mandibulae anterior superficialis (lmasf), m. levator mandibulae anterior articularis (lmaa) и другая порция lmpp, совместно приводящие нижнюю челюсть и поворачивающие передние концы меккелевых хрящей
Возможно, таким же образом произошло расширение функций жаберных дуг позвоночных при возникновении челюстноротых. Связанное с интенсификацией дыхания, т. е. с усилением тока воды, выталкиваемой из ротоглоточной полости через жаберные щели и омывающей жабры, увеличение подвижности жаберных дуг в ряду позвоночных достигается двумя способами. У бесчелюстных - истончением и увеличением эластичности не расчлененных дуг (круглоротые), у челюстноротых - расчленением жаберных дуг на подвижные друг относительно друга элементы. Расчлененная жаберная дуга при сжатии глотки, выталкивающем воду через жаберные щели, складывается, что обеспечивает большую амплитуду движения. Однако при этом дуга оказывается способной зажимать в глотке комки пищи, не давая им выноситься с током воды через жаберные щели или засорять их. Видимо, на этой основе третья жаберная дуга превратилась в челюсти, тогда как две передние редуцировались, освобождая место для ее увеличения.
Итак, в основе самой возможности функциональных преобразований организма в ходе филогенеза лежат два принципа: мультифункциональность и множественное обеспечение функций. Чем больше функций выполняет данная система, тем в большем числе направлений она может меняться в ходе последующего филогенеза, т. е. тем выше ее эволюционная пластичность. Аналогично, чем больше систем участвует в обеспечении той или иной биологически важной функции, тем шире возможности преобразования любой из этих систем, причем сама функция при этом сохраняется. Типы функциональных преобразований организации многочисленны, И количество их, по-видимому, будет увеличиваться, так как введение нового эволюционного принципа почетно для его автора. Тип, по которому на данном отрезке филогенеза осуществляется преобразование той или иной структуры, определяется несколькими параметрами: направлением отбора по данной функции, структурой, которая данную функцию выполняет (включая сюда функциональные и пространственные связи этой структуры с другими), а также функциональной ее нагруженностью (Креславский, 1977), т. е. важностью для организма выполняемых ею функций. Все эти параметры, взаимодействуя между собой, в конкретных эволюционных ситуациях приводят к тому, что каждый конкретный тип филогенетических преобразований органов описывает лишь отдельные стороны процесса. Реальный процесс филогенеза органов может быть более или менее адекватно охарактеризован лишь комплексом типов, а реакция организма на действие того или иного направления отбора не может быть сведена к изменениям отдельных структур и их функций. Даже при отборе тю одной конкретной функции перестраивается вся система функций данной структуры. Поэтому трудно ожидать, что описание новых принципов и типов изменений органов в связи с их функцией будет способствовать реальному прогрессу в разработке проблем функциональной эволюции.
Структура допускает осуществление ряда функций, но каждая функция осуществляется той или иной структурой. Поэтому можно утверждать, что на каждом этапе филогенеза структура определяет направление и саму возможность дальнейшего преобразования самой себя в ходе филогенеза. Сложившаяся организация наряду с условиями окружающей среды и во взаимодействии с ними направляет дальнейший ход филогенеза, причем чем пластичнее организация, тем шире спектр возможных ее изменений при изменении экологической обстановки.
Пластичность эколого-физиологических реакций организма, а отсюда и эволюционная пластичность во многом определяются характером связи между структурой и функцией. Чем выше функциональная нагрузка на признак, тем жестче связь между структурой и выполняемой ею функцией. Во всяком случае, чем выше функциональная нагрузка на признак, тем слабее его индивидуальная изменчивость (Креславский, 1977). Функциональная нагрузка, определяемая значением данной функции для организма, возрастает при интенсификации функций и поддерживается стабилизирующим отбором. Чем выше функциональная нагрузка, тем меньше эволюционная пластичность органа и тем ортогенетичнее (Гиляров, 1976) его дальнейшая эволюция, если она все еще возможна на данном этапе филогенеза.
Снижение мультифункциональности биологически важных систем, особенно если их функции связывают организм со средой, ведет к ограничению эволюционной пластичности таксона и чревато вымиранием его при резких изменениях среды. Видимо, одно из значений множественного обеспечения биологически важных функций заключается в снижении функциональной нагрузки на каждую из участвующих в обеспечении функций структур и сохранении (повышении) их пластичности. Другим путем обхода этого ограничения является комбинирование блоков (Уголев, 1985), которое усилилось с возникновением многоклеточности и явилось, по всей вероятности, важным фактором поддержания целостности и в то же время повышения дифференцированности организма по мере филогенеза.
Чем слабее связь между структурой и функцией, ею осуществляемой, тем выше эволюционная пластичность структуры. Это определяется двумя параметрами: 1) при ослаблении связи структуры и функции изменения структуры меньше сказываются на осуществлении функции; 2) при менее жесткой связи структуры и функций, ею выполняемых, структура может быть более мультифункционалыюй. Однако, как отмечено выше, повышение мультифункциональности приводит к ухудшению осуществления главной функции и к снижению адаптивности системы, а интенсификация функций, связанная с возрастанием функциональной нагрузки на признак, связана с уменьшением числа функций и со снижением эволюционной пластичности структуры.
Та или иная из двух противоречивых тенденций (ослабление или усиление связи между структурой и функцией) в ходе филогенеза каждой конкретной системы побеждает в зависимости от конкретной ситуации. Но поскольку они обе важны, наиболее общим способом разрешения их противоречия в ходе прогрессивной эволюции оказалось возникновение иерархии функций. Чем выше иерархический уровень данной функции, тем большее число систем участвует в ее обеспечении и тем больше автономность данной функции по отношению к любой из систем, ее обеспечивающих.
Наиболее автономны, а потому лабильны такие сложные функции, как поведение. Не случайно именно поведенческие реакции в первую очередь определяют адаптацию высших форм жизни и возможность их дальнейшей эволюции. Таким образом, организация особи сама по себе ограничивает число возможных направлений ее преобразования в ходе дальнейшей эволюции. Чем выше функциональная нагрузка на признак, тем ниже его мультифункциональность и тем меньшее число направлений дальнейшей эволюции этого признака. Вторым механизмом снижения мультифункциональности является усиление функциональных связей внутри системы (см. 2.4), приводящее к формированию блоков. Сохранение и повышение эволюционной пластичности достигается несколькими путями: комбинированием блоков, множественным обеспечением функций и иерархичностью организации морфофункциональных систем, чему во многом способствует комбинирование блоков.
Таким образом, целесообразно рассматривать три аспекта функциональной эволюции систем I типа: прогрессивное развитие данной морфологической структуры, ее редукцию и ее перестройку. Системы II типа (системы множественного обеспечения биологически важных функций) могут рассматриваться как состоящие из некоего множества систем первого типа. Системы III типа более сложны, так как включают непосредственные функциональные взаимодействия между компонентами. Эволюция систем II и III типов рассмотрена ниже (см. 2.4). Здесь уместно еще раз подчеркнуть, что число функций, выполняемых данной системой в данный момент времени, жестко детерминирует число потенциально возможных направлений ее эволюции. Реализация того или иного из этих направлений детерминируется па микроэволюционном уровне, т. е. зависит от направления естественного отбора. Взаимодействие подсистем (а все морфофункциональные системы организма, по сути, представляют собой системы III типа, и выделение систем I и II типов является более или менее сильным упрощением реальной ситуации) сужает число потенциально возможных направлений эволюции и еще более жестко канализирует ее.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'