2.4.3. Перестройка координации
Поскольку координации ограничивают число возможных направлений эволюции организма и чем прочнее координационные связи, тем жестче эти ограничения, следует рассмотреть вопрос о прочности координаций, причинах их перестройки или исчезновения. Согласно И. И. Шмальгаузену, наименее прочны биологические координации, тогда как топографические и динамические более устойчивы. Действительно, при смене адаптивных зон (см. 2.5) биологические координации легко разрываются и заменяются новыми. Так, у наземных млекопитающих, в том числе и у насекомоядных, развитие задних (толчковых) коtvgнечностей координировано с развитием передних конечностей таким образом, что задние конечности всегда сильнее. Напротив, у грызунов, ведущих подземный образ жизни (цокоров, слепышей, слепушонок), и особенно у кротов, соотношение направлений эволюций конечностей меняется: преимущественное развитие получают передние конечности, что связано либо с выталкиванием земли из ходов при опоре на передние конечности (Гамбарян, 1960), либо непосредственно с рытьем (кроты). Биологические координации, видимо, могут ограничивать число возможных направлений эволюции, входящих в данную координацию органов лишь до тех пор, пока таксон эволюционирует в данной адаптивной зоне.
Топографические и динамические координации разрываются или перестраиваются, по-видимому, действительно реже, чем биологические. Однако трудно судить о том, является ли их большая стабильность следствием прочности связей внутри координационной системы или следствием более универсального значения этих связей. Топографические координации обусловлены планом строения (или его частными проявлениями), т. е. онтогенетическими механизмами и стабильны именно благодаря важности этих механизмов. Однако можно показать, что такие координации, как соотношение расположения поясов конечностей, висцерального скелета относительно осевого черепа и т. п., перестраиваются при смене адаптивных зон не менее пластично, чем биологические координации.
Действительно, у морского черта, своеобразной придонной рыбы, брюшные плавники оказываются впереди грудных (Северцов, 1939); у головастиков бесхвостых амфибий задний край подъязычного жаберного аппарата смещается вперед относительно ушной капсулы по сравнению с хвостатыми амфибиями (Северцов, 1974; Edgeworth, 1935). Как известно, у скатов жаберные щели сместились на брюшную сторону тела, а брызгальце, передняя жаберная щель - на дорсальную. Иными словами, топографические координации оказываются не более стабильными, чем биологические, и также перестраиваются при изменении среды обитания. Представления о большей устойчивости топографических координаций, видимо, могут быть обусловлены тем, что такие соотношения, как план строения первичноротых или вторичноротых, билатеральность симметрии, расположение мозга в черепно-мозговой полости и т. п. пространственные соотношения, как правило, сохраняются в течение длительных периодов филогенеза независимо от смены среды обитания в ходе филогенеза. Нетрудно, однако, показать, что неизменность пространственных соотношений типа плана строения или симметрии обусловлена не прочностью этих соотношений самих по себе, а прочностью морфогенетических механизмов, их формирующих, которая в свою очередь обусловлена необходимостью поддержания целостности организма в онтогенезе. Общеизвестно, околь глубоки перестройки топографии внутренних органов и, в частности, нарушения симметрии, вызванные переходом к ползанию у змей.
Динамические координации, насколько об этом можно судить, действительно, более прочны, чем биологические и топографические. Во всяком случае, функциональные связи внутри организма, как правило, сохраняются при смене адаптивных зон, как бы глубоко при этом ни менялись компоненты данной координации (функциональной системы). Такие координации, как связь глаз и зрительных центров мозга, скелетных элементов и управляющих ими мышц, отделов пищеварительного тракта и т. д. и т. п., обычно сохраняются независимо от смены среды обитания до тех пор, покуда существуют составляющие данную координацию компоненты, и при редукции исчезают совместно.
Мало того, в ряде случаев именно динамические координации столь жестко ограничивают число возможных направлений преобразования организации входящих в них органов, что приобретают характер "организационных запретов эволюции" (Иорданский, 1985; Тимофеев-Ресовский, Воронцов, Яблоков, 1969). Примером подобного "запрета" может служить координация дыхательных поверхностей у амфибий, ранее рассмотренная вкратце в качестве примера множественного обеспечения биологически важных функций. Все три дыхательные поверхности современных амфибий (легкие, кожа и слизистая ротоглоточной полости) функционировали у предков наземных позвоночных - кистеперых рыб. В условиях бедных кислородом прогреваемых водоемов, в которых обитали Rhipidistia (Шмальгаузен, 1964), ротоглоточно-легочный комплекс имел значение дополнительного (к жабрам) источника получения кислорода из атмосферы, а кожа служила дополнительной поверхностью выведения из организма углекислоты. Такое разделение функций было обусловлено тем, что воздух захватывался в ротоглоточную полость и легкие при всплывании рыбы, а выведение углекислоты через легкие привело бы к повышению частоты всплывания рыбы за свежим воздухом, т. е. излишне демаскировало бы этих подстерегающих хищников (Шмальгаузен, 1964).
Редукция жабр, происходившая в процессе освоения суши, привела к компенсаторному усилению газообмена через все три дыхательные поверхности, причем кожа приобрела значение органа, эффективно участвующего в потреблении кислорода. Однако амфибии, как и их предки - кистеперые, продолжают использовать для вентиляции легких нагнетательный насос - вертикальные колебания дна ротоглоточной полости, основанные на поворотах подъязычного аппарата. Сохранение нагнетательного насоса вентиляции легких совокупно с интенсификацией кожного газообмена обусловили, с одной стороны, редукцию ребер, а с другой - сохранение в течение всего филогенеза амфибий именно трех дыхательных поверхностей: ни одна из них не способна полностью обеспечить газообмен животного во всем диапазоне колебаний внешних условий (см. выше). Исключение составляют лишь безлегочные виды Urodela, у которых редукция легких была связана с интенсификацией кожного дыхания и совершенствованием механизма выбрасывания языка. Нетрудно заметить, что множественное обеспечение газообмена амфибий, которое следует рассматривать как динамическую координацию (баланс) трех дыхательных поверхностей, резко ограничивает эврибионтность амфибий. Необходимость кожного газообмена делает неизбежной проницаемость кожи для воды и существование амфибий лишь во влажных биотопах.
Таким образом, устойчивость динамической координации ограничивает степень дивергенции. В отличие от биологических и то-пографических координаций, достаточно легко разрывающихся при смене сред обитания, динамические могут запрещать саму смену сред. Это явление заставляет проанализировать два вопроса: 1. При каких условиях динамическая координация не может быть разорвана? 2. Каковы условия разрыва динамических координаций?
Очевидно, у амфибий при интенсификации легочного дыхания, как это произошло у рептилий на основе развития разрежающего насоса, кожное дыхание могло бы редуцироваться и ограничение на освоение суши для амфибий было бы снято. Иными словами, баланс трех дыхательных поверхностей амфибий обусловлен несовершенством нагнетательного насоса вентиляции легких, позволяющим использовать лишь смешанный воздух (Северцов, Соколов, 1972, 1975; Krogh, 1904) и сдерживающим эволюцию структуры легких. Лишь у вторичноводных Pipidae легкие приобретают сложную ячеистую структуру (de Jongh, 1972), что сопряжено с очень мощным развитием подъязычного аппарата. У Pipidae прогрессивное развитие подъязычного аппарата (см. рис. 19) коррелировано с редукцией языка. У остальных Anura язык связан с подъязычным аппаратом мышцей-ретрактором - М. hyoglossus. Прикрепляется язык к ментальным косточкам нижней челюсти, а выбрасывается на основе сложного взаимодействия мышц и лимфатической полости - sinus sublingualis (см. рис. 11, 16) (Северцов, 1974).
У Urodela, которым также свойственно выбрасывание языка (см. рис. 11), этот орган сохранил первичную для Tetrapoda и более тесную, чем у Anura, связь с подъязычным аппаратом (см. рис. 12, 13). Поскольку эта связь и возникла в филогенезе для обеспечения подвижности языка (Северцов, 1974), ее надо рассматривать в качестве динамической, а не топографической координации. У всех современных Urodela язык выбрасывается благодаря горизонтальным движениям подъязычного аппарата, которые тем самым обеспечивают питание этих животных. Таким образом, у Anura разобщение механизмов выбрасывания языка и вентиляции легких стимулировало развитие легких и уменьшение роли кожного дыхания. У Urodela этого не произошло. Разобщение механизмов захватывания пищи и вентиляции легких для этих животных равноценно либо редукции языка, т. е. коренной перестройке способа питания, либо развитию механизма движений дна ротоглоточной полости без участия подъязычного аппарата, что тоже невозможно. Иными словами, разрыв динамической координации механизмов дыхания и захватывания пищи повлек бы за собой дальнейшее снижение приспособленности хвостатых амфибий. Именно поэтому подобный разрыв невозможен, и именно поэтому эволюция дыхания Urodela пошла по пути интенсификации функции кожного дыхания и ослабления функции легочного. Urodela - единственная группа наземных позвоночных, в пределах которой имеются безлегочные формы. Ослабление функции легочного дыхания, в крайней форме выражающееся в редукции легких, привело к биологическому регрессу хвостатых амфибий. Современная фауна Anura представлена приблизительно 3300 видами. Urodela насчитывается около 300 видов, объединяемых в 62 рода. Представители примерно 20 родов ведут вторично водный образ жизни (в том числе и неотеничные формы), а 26 родов (42%) - монотипические. Данные по числу видов несколько занижены, но соотношение может быть изменено лишь в пользу Anura (Ананьева и др., 1988).
Из вышеизложенного следует, что невозможность разрыва динамической координации механизмов дыхания и захватывания пищи привела через сохранение координации трех дыхательных поверхностей к снижению эволюционной пластичности Urodela, еще более глубокому, чем это наблюдается у Anura. Само же сохранение динамической координации механизмов вентиляции легких и ловли добычи обусловлено тем, что перестройка любого из них означает снижение приспособленности либо по питанию, либо по газообмену. Таким образом, "организационный запрет" направления эволюции представляет собой ситуацию, когда разрыв или перестройка координационной системы невозможна из-за того, что подобная перестройка привела бы к снижению приспособленности. Следовательно, разрыв динамической координации возможен только в том случае, если он не приводит к снижению приспособленности.
По-видимому, наиболее прочны динамические координации на глубоких (прежде всего молекулярном) уровнях организации, рассматриваемые А. М. Уголевым (1985) как функциональные блоки. Эти динамические координации могут перестраиваться, вероятно, только при условии ослабления функциональных связей внутри или между ними. Подобное ослабление связей может происходить прежде всего при изменении условий функционирования блока, что Достигается перекомбинацией или транспозицией блоков. С этих позиций А. М. Уголевым (1985) рассмотрено происхождение ядов Животных. Яды скорпионов, насекомых, рыб, амфибий и рептилий представляют собой не что иное, как энзимы, в норме служащие регуляторами различных физиологических функций животных. Так, яды, выделяемые кожными железами амфибий (например, бомбезин), представляют собой релизинг-факторы позвоночных. Бомбезин обнаружен в стенке кишки и в ЦНС млекопитающих (Уголев, 1985). Пресинаптические нейротоксины змей, таких как кобра, австралийская тигровая змея (Notechis scuatatus), тайпан (Oxiuranus scutellatus) и др., по своей аминокислотной последовательности высоко гомологичны с основной фосфолипазой из поджелудочной железы и слюнных желез млекопитающих (Eaker et al., 1976). Ингибиторы протеаз из ядов Viperidae и Elapidae высоко гомологичны панкреатическому основному ингибитору трипсина (Iwanaga et al., 1976). Выделение энзимов, перенесенное вовне организма, привело к смене функций, превратив их из регуляторов метаболизма в яды. Это еще один неизвестный до работ А. М. Уголева способ смены функций, связанный не с отбором по второстепенным функциям, а с сохранением главной функции в новых условиях функционирования.
Транспозиция подобных блоков, высвобождая функциональные связи, т. е. ослабляя координацию, приводит к дальнейшей эволюции блоков. Токсины ядов животных лишь высокогомологичны, но не тождественны энзимам-регуляторам, действующим внутри организма. Молекуляоная структура их эволюционировала. Так, известны два типа нейротоксинов змеиных ядов, связывающихся с ацетилхолиновыми рецепторами. Один тип представлен токсинами, состоящими из 60-62 аминокислотных остатков с 4 дисульфидными мостиками; другой - из 71-74 аминокислотных остатков с 5 дисульфидными мостиками. В ядах животных присутствует смесь сходных по структуре и физиологическому действию токсинов. В то же время яд каждого вида представлен более чем одним типом токсинов, более чем одним блоком, по терминологии Уголева. Например, яд кобры включает пресинаптические нейротоксины, постсинаптические нейротоксины, кардиотоксины и протеазы. Это свойство ядов - их многокомпонентность, по-видимому, универсально для животных (Bucherl, Buckley, 1971; Hashimoto, 1979; Lin Sian et al., 1976; Maeda et al., 1976; Somejima et al., 1976; Watanabe et al., 1976). Видимо, отбор контролирует не столько токсичность каждого из индивидуальных токсинов, сколько эффективность всей смеси - яда в целом. Возможно, разнообразие блоков, входящих в яд данного вида, обеспечивает более широкий спектр жертв, тогда как некоторое снижение токсичности тех или иных компонентов может быть компенсировано увеличением продукции яда.
Не рассматривая генетические механизмы формирования полиморфизма токсинов, необходимо отметить, что сохранение высокой степени гомологии каждого из них нормальным энзимам жертвы - непременное условие эффективности яда. Иными словами, яды становятся ядами не только потому, что имеются готовые блоки, обеспечивающие их продукцию, но и потому, что имеются готовые блоки, обеспечивающие их рецепцию. Таким образом, ослабление функциональных связей данного блока ведет к его дестабилизации и дальнейшей эволюции. Это, по-видимому, обязательное условие перестройки высококоординированных функциональных систем.
Можно указать два не связанных с транспозицией механизма, обеспечивающих разрыв динамических координаций без снижения приспособленности. Либо данная система должна быть "запараллелена" (множественное обеспечение функций), тогда усиление одной из "параллелей" может создать возможность для разрыва другой, либо разрыв координации, приводящий к снижению приспособленности на данной стадии онтогенеза, компенсируется повышением приспособленности другой стадии онтогенеза, возникающим в результате разрыва данной координации. Первый случай иллюстрируется данными Н. С. Лебедкиной (1980). Как отмечено выше, у кистеперых рыб praemaxillare укреплялась главным образом на массивном эндокране, но одновременно соединялась и с покровными костями крыши черепа. В частности, у Osteolepiformes она сливалась с передней nasale. С выходом по-звоночных на сушу это второе соединение приобрело решающее значение. В результате связь praemaxillare с эндокраном потеряла функциональное значение, координация была разорвана, и эндокран редуцировался.
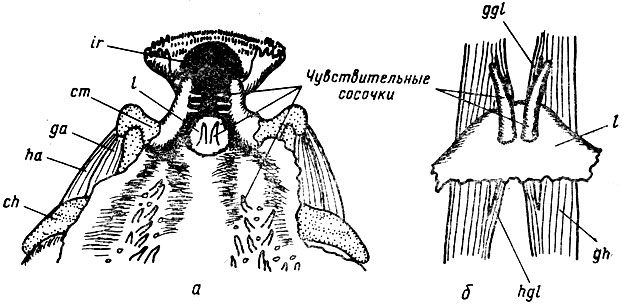
Рис. 22. Передняя часть ротоглоточной полости (а) и язык (б) головастика Pelobates fuscus: ch - ceratohyale, cm - cartilago mecceli, gh - m. geniohyoideus, ggl - m. genioglossus, ha - m. hyoangularis, ir - infrarostrale, I - язык, qa - m. quadratcangularis
Второй способ можно проиллюстрировать разобщением меха-низмов дыхания и ловли добычи у бесхвостых амфибий. У Anura в отличие от Urodela дефинитивный язык формируется не в процессе метаморфоза, а на ранних этапах личиночного развития. Это обусловлено необходимостью контроля за током воды, поступающим в ротовую полость и несущим частички пищи, откушенные роговым "клювом". На языке личинок Anura расположены два длинных сосочка с вкусовыми почками, к основаниям которых прикрепляются мышцы языка (рис. 22). Поскольку язык развивается именно у личинок, он утрачивает связь с подъязычным скелетом и в процессе метаморфоза приобретает более тесную связь с симфизом нижней челюсти. Разобщение механизмов выбрасывания языка и вентиляции легких у Anura в свою очередь обусловило возможность прогрессивного развития обоих этих механизмов (см. выше). Редукции легких у бесхвостых амфибий не наблюдается. Описанная выше динамическая координация разрывается.
Таким образом, координации играют в филогенезе двоякую роль: они ограничивают число возможных направлений эволюции организма и одновременно повышают скорость его адаптации в процессе отбора. Адаптивная роль координаций очевидна: они обусловливают целостность организма в ходе филогенетических преобразований.
Подводя итоги анализу функциональных преобразований организации особи в ходе филогенеза, т. е. проблеме функциональной дифференциации организма и его целостности в ходе этой дифференциации, необходимо подчеркнуть, что в основе их лежат два основных требования: необходимость перестраиваться в соответствии с изменениями внешней среды и необходимость поддерживать гомеостаз организма в ходе этих, подчас весьма глубоких и разнонаправленных перестроек. Сложности усугубляются тем, что организм вынужден адаптироваться в условиях ограниченности ресурсов среды, приводящих к тому, что любая вновь возникающая адаптация приводит к хотя бы частичной утрате приспособленности по другим признакам, которая может быть компенсирована лишь дальнейшим увеличением мультифункциональности и координированности частей организма. И то и другое повышает экономичность организации, так как снижает число структур, специально определяющих те или иные функции и пластичность организации. Видимо, сама мультифункциональность обусловлена двумя причинами: невозможностью создать специализированные системы для каждой функции, как бы они ни были адаптивны, и поддержанием целостности организации - взаимосвязи частей организма осуществляются через их функции.
Вторым существенным ограничением свободы преобразований частей организма является невозможность одновременного выполнения данной мультифункциональной системой более чем одной активной функции. Невозможность "делать два дела сразу" была показана еще A. H. Северцовым (1939) и подчеркнута С. П. Масловым (1980). Действительно, если данная структура выполняет в данный момент времени какую-либо функцию, в то время как другая функция, необходимая в тот же момент времени для поддержания гомеостаза организма, ею не выполняется из-за указанного ограничения, то при множественном обеспечении функций это ограничение снимается.
Следствием этого положения является необоснованность попыток выявления в организме "лишних деталей". Например, указания на избыточность нейронов ЦНС или наличие избыточной ДНК. Если те или иные компоненты организма при выявлении их главной функции оказываются (в эксперименте) как бы лишними, не выполняющими данную функцию, означает что они выполняют в данный момент времени другие функции, а не их избыточность (ср. Федоров, 1988). Как отмечено выше, избыточность функционирования, но не структуры, существует, однако она определяется адаптацией к высоким функциональным нагрузкам.
Непонимание этого, как справедливо отметил С. П. Маслов (1980), "приводит в недоумение" последователей теории номогенеза (ср. Мейен, 1979). Однако сложность взаимодействий внутри организма, обусловливая и его гомеостаз, и его эволюционную пластичность, одновременно служит важнейшим фактором, направляющим эволюцию. Сколь бы широко ни была индивидуальная изменчивость, в ходе эволюции могут быть отобраны лишь такие изменения, которые не снижают ни эффективности функционирования, ни целостности организации, причем адаптивный эффект наблюдаемых преобразований должен перекрывать вызываемое данным преобразованием снижение приспособленности по другим признакам.
Таким образом, мозаичность организации отражает прежде всего пластичность ее. Пока системы координаций не сформировались, каждая часть организма обладает большей автономностью и большим числом степеней свободы для ее эволюционных преобразований. По мере формирования координационных систем число связей любой данной системы с другими частями организма растет, сами эти связи становятся более прочными и число возможных направлений эволюции данной системы сокращается - формируется структурный тип данного таксона. В результате пластичность организации целостного организма уменьшается в том смысле, что сокращается число возможных направлений его эволюции. Вместе с тем благодаря возрастанию интегрированности организма все его преобразования проходят быстрее и более координирование.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'