2.5.4. Монофилия и полифилия в происхождении таксонов надвидового ранга
Проблема монофилии и полифилии в происхождении надвидовых таксонов - одна из наиболее дискутируемых проблем теории филогенеза. Вызваны эти дискуссии тем, что точное выяснение происхождения крупных таксонов всегда затруднено, а широкое распространение параллелизмов при ограниченности числа пригодных для анализа признаков часто не допускает однозначного толкования выявляемых филогенетических связей. Это порождает возможность как полифилетической, так и монофилетической интерпретации соотношений анцестральных и дочерних систематических групп.
Монофилия, в строгом значении этого термина, подразумевает, что весь дочерний таксон независимо от его ранга и объема возник от одного исходного вида. И. И. Шмальгаузен (1969) специально подчеркивал, что в качестве исходного следует рассматривать именно вид, а не особь или пару особей. Это требование вытекает из представлений о популяции как элементарной единице эволюционного процесса. Оно породило представление, что вид - родоначальник таксона должен входить в состав этого, дочернего, таксона (Очев, 1966 и др.). Такой подход позволяет рассматривать всю дивергенцию дочернего таксона как результат преобразования организации потомков этого вида, а сам таксон считать естественным (Расницын, 1969; Татаринов, 1976; Wagner, 1968).
Под полифилией обычно понимают возникновение исходного таксона от нескольких или многих исходных систематических групп. Признание полифилии в качестве единственного пути эволюции приводит к представлению об уменьшении разнообразия жизни в ходе эволюции (Татаринов, 1976). Действительно, если каждый таксон возникает более чем от одного исходного, то со временем число таксонов любого ранга уменьшается. Такой подход подразумевает, что эволюция идет только путем конвергенции. Уменьшение разнообразия жизни будет происходить и в том случае, если считать, что число систематических групп анцестрального таксона равно числу групп дочернего таксона. Эта точка зрения, соответствующая теории ортогенеза, в сочетании с вымиранием таксонов, происходившим на протяжении всей истории биосферы, также предполагает оскудение жизни по мере ее эволюции. По сути дела, обе теории финалистичны. Обе они, подразумевая отрицание дивергенции, представляют в настоящее время лишь исторический интерес, хотя А. Н. Северцов (1939) и считал, что лишь две крайние точки зрения - ортогенез и строгая монофилия - не содержат внутренних противоречий и имеют право на существование. Согласно его рассуждениям, признание монофилии в происхождении одного таксона означает невозможность отрицания такого же происхождения любого другого таксона. Поскольку дивергенция доказана, а ортогенез с позиций дарвинизма невероятен, справедлива теория монофилии. Нетрудно заметить, что приведенное рассуждение А. Н. Северцова отражает существовавшую в то время крайнюю недостаточность конкретных данных о филогенезе надвидовых таксонов. С накоплением таких данных этот подход утратил актуальность.
Современные сторонники теории полифилии не отрицают дивергенцию как явление. Вопрос ставится в другой плоскости: могут ли два или большее число независимых видов пересечь демаркационную линию, разграничивающую исходный и дочерний таксоны. Обсуждение этой проблемы наряду с обсуждением теории полифилии собственно и теории строгой монофилии привело к разработке компромиссного подхода: теории широкой монофилии (Simpson, 1959, 1961).
Примером собственно полифилетических представлений могут служить взгляды Е. Стеншио (Stensio, 1963) на филогенез круглоротых: миноги возникли от Pteraspides - подотряда Ostracodermi, а миксины от Cephalaspides - другого надотряда того же класса. Не менее известна теория полифилетического происхождения наземных позвоночных Э. Ярвика (Jarvik, 1952-1972). Согласно этой теории лепоспондильные стегоцефалы и их потомки - хвостатые амфибии - возникли от одной группы кистеперых рыб - Роrоlepiformes, тогда как рахитомные стегоцефалы, бесхвостые амфибии, рептилии и млекопитающие появились независимо друг от друга от другой группы кистеперых рыб - Osteolepiformes (Jarvik, 1967). Лишь птицы, согласно этой теории, произошли от примитивных рептилий, а не непосредственно от кистеперых рыб. В то же время сами Osteolepiformes и Porolepiformes дивергировали от общего предка.
Компромиссная концепция широкой монофилии, развитая Дж. Симпсоном (Simpson, 1959, 1961) и поддержанная в отечественной литературе Н. В. Тимофеевым-Ресовским, Н. Н. Воронцовым и А. В. Яблоновым (1969), сводится к утверждению, что таксон следует считать монофилетическим в том случае, если его ранг выше или равен рангу анцестрального таксона независимо от числа реальных филогенетических связей между обеими группами. Блестящая критика этой концепции дана Л. П. Татариновым (1976). Действительно, в зависимости от ранга исходного и дочернего таксонов один и тот же тип филогенеза можно, пользуясь подходом Дж. Симпсона, представить и как монофилию и как полифилию. Так, с позиций концепции широкой монофилии изложенные взгляды Э. Ярвика на происхождение наземных позвоночных при рассмотрении Tetrapoda в целом (надкласс) представляются монофилией, но по отношению к амфибиям (класс) - первой группе наземных позвоночных - полифилией. Мало того, поскольку в разных системах ранг даже хорошо очерченных групп варьируется, меняя этот ранг, несложно, в рамках концепции Дж. Симпсона, любую филогенетическую схему представить и как монофилетическую и как полифилетическую. Так, при рассмотрении Osteopiformes и Porolepiformes в качестве отрядов подкласса Phipidistia (Воробьева, Обручев, 1964) возникновение от них класса амфибий, с позиций Дж. Симпсона, выглядит как монофилия, но при рассмотрении этих же групп в ранге классов надкласса кистеперых рыб (Stensio, 1963) происхождение класса амфибий должно быть признано полифилетическим.
Таким образом, концепция широкой монофилии, или, по терминологии Г. Смита (Smith, 1967), макромонофилии, не позволяет объективно интерпретировать филогенетические связи таксонов. Л. П. Татаринов (1976) настаивал на строгой интерпретации палеонтологических данных. Когда палеонтологический материал достаточно полон для того, чтобы обсуждать происхождение больших таксонов на уровне родов или семейств, обычно выявляются две или больше систематические группы исходного таксона, каждая из которых с успехом может считаться анцестральной для всего дочернего таксона, причем ни для одной из этих групп не удается доказать, что именно она представляет собой истинную предковую группировку. В признании более или менее одновременного пересечения границы между исходным и дочерним таксонами несколькими морфологически близкими группами и состоит суть концепции Татаринова.
Например, в настоящее время известно около 30 видов высших териодонтов, "почти достигших маммального уровня организации" (Татаринов, 1976, с. 207). В то же время ряд видов примитивных мезозойских млекопитающих еще не имели полного набора диагностических признаков этого класса, в частности, у них сохранились постдентальные кости нижней челюсти. Л. П. Татаринов считает невероятным, чтобы границу между териодонтами и млекопитающими пересек лишь один единственный из нескольких десятков готовых к такому переходу видов. Кроме того, этот же автор совершенно справедливо отметил, что строгая монофилия ни в одном случае не доказана на конкретном материале.
Из сказанного следует, что выбор между широкой полифилией и ограниченной полифилией, с одной стороны, и ограниченной полифилией и строгой монофилией, с другой, возможен только на основе построения теоретических моделей. Фактических данных для решения этого вопроса, как и в 30-х гг., когда А. Н. Северцов обсуждал эту проблему, все еще недостаточно.
Признание полифилии в сочетании с признанием дивергенции приводит к признанию возможности полного слияния прежде самостоятельных таксонов и к расширению концепции сетчатого родства (Шмальгаузен, 1969) на надвидовые таксоны. Возникший на основе полифилии таксон в процессе освоения занятой им адаптивной зоны должен испытывать дивергенцию (адаптивную радиацию). В ходе адаптивной радиации нарастает число дочерних систематических групп, которые в разных сочетаниях станут родоначальниками следующего "поколения" полифилетических таксонов. Иными словами, сочетание дивергенции и полифилии дает картину сетчатой эволюции в том виде, как ее нарисовал Т. Добжанский (Dobzhansky, 1951), для взаимодействия эволюционирующих популяций внутри вида. Такая интерпретация полифилии избавляет от признания ортогенеза и (или) уменьшения числа видов в ходе эволюции, но оставляет открытым вопрос о возможности слияния воедино, вплоть до полной неразличимости, нескольких исходных таксономических групп.
Экстраполяции концепции сетчатой эволюции довидового уровня включительно сделана И. С. Даревским (1974). Она основана на случаях гибридогенного видообразоваия, показанного для скальных ящериц (Даревский, 1974), для ряда костистых рыб (Васильев, 1985) и для ряда аллополиплоидов у растений. Однако гибридогенное видообразование едва ли можно интерпретировать как сетчатую эволюцию. Если два близких вида, геномы которых еще не утратили совместимости, при снятии пространственных или биотопических изоляционных барьеров, через партеногенез или без него могут на основе гибридизации дать начало новому, третьему виду, адаптированному к нарушенным биогеоценозам, то гибридогенез этого дочернего вида с каким-либо другим видом, имеющим гибридогенное происхождение, - событие практически невероятное: Отбор на изоляцию будет действовать тем интенсивнее, чем сильнее гибридизация. Репродуктивная изоляция - один из основных критериев вида - и межвидовая конкуренция будут препятствовать сетчатой эволюции на видовом уровне. Именно поэтому гибридогенное видообразование - относительно редкий случай, исключение, которое подтверждает правило: видообразование идет через внутривидовую дифференциацию и обособление подвидов.
Гибридизация, т. е. слияние в единую популяцию представителей систематических групп надвидового ранга, с позиций генетики невероятна. По мере дивергенции генетическая обособленность, а отсюда и несовместимость геномов дивергирующих групп нарастают, и уже на уровне хорошо обособившихся видов одного рода даже искусственное скрещивание (если оно и удается) часто приводит к понижению жизнеспособности или к снижению плодовитости потомства. Поэтому невозможно представить себе сетчатую эволюцию, а тем самым и истинную полифилию, т. е. возникновение единого в генетическом смысле таксона на основе нескольких анцестральных групп. Однако поскольку при исследовании филогенеза о родстве судят по сходству, то полифилия может быть интерпретирована как возникновение от нескольких исходных форм такого таксона, который не обладал бы генетическим и филогенетическим единством, но на основе всех доступных методов сравнительного исследования представлялся бы единым. При такой интерпретации вопрос о полифилии и монофилии из плоскости теории филогенеза переходит в плоскость теории систематики.
Действительно, при условии доказательства возможности происхождения надвидового таксона от двух или нескольких исходных групп независимо от степени близости этих групп в отсутствие возможности доказательства обратного возникает вопрос уже не о филогенетических связях дочернего таксона, а о том, какие выводы должен сделать систематик при установлении полифилии. Таким образом, проблема полифилии и монофилии в происхождении надвидовых таксонов сводится к решению следующих вопросов:
1. Могут ли два или большее число таксономических групп независимо друг от друга начать осваивать новую адаптивную зону?
2. Возможна ли ситуация, в которой потомки этих таксономических групп будут настолько сходны между собой, чтобы на основе всех доступных методов сравнительного исследования можно было бы считать их единой таксономической группой?
3. К каким выводам должен прийти систематик при установлении факта полифилетического происхождения таксона?
Прежде чем перейти к обсуждению этих вопросов, а их решение может быть основано лишь на построении теоретической модели, использующей сведения о микроэволюции, следует еще раз обсудить вопрос о том, какую группу считать родоначальницей таксона. Если вслед за В. Г. Очевым (1966) признать в качестве таковой вид, входящий в состав дочерней группы, т. е. уже освоивший новую адаптивную зону, то теряется смысл установления филогенетического родства исходной и дочерней групп. Даже при гибридогенном видообразовании сначала возникает гибридная популяция, а затем подчас очень сложными путями (Васильев, 1985; Даревский, 1974) на ее основе - новый вид. Если мы будем считать эту популяцию анцестральной, то гибридогенное видообразование нельзя считать полифилетическим: вид возникает из единой популяции. Однако при таком подходе утрачивается и представление о происхождении этого вида от видов-предков. На практике при обсуждении полифилии или монофилии и видов и таксонов надвидового ранга, как и в любых случаях установления филогенетического родства, в качестве предков рассматривают формы, принадлежащие к исходному таксону: для тетрапод - это кистеперые рыбы, для млекопитающих - териодонты и т. д. Иными словами, предложение В. Г. Очева (1966) неприемлемо.
При обсуждении проблемы монофилии и полифилии необходимо исследовать родственные связи между двумя таксонами, а не внутри одного из них. На практике это выражается в сравнении групп: если сходство между таксономическими группами, составляющими дочерний таксон, выше, чем между каждой из этих групп и сравниваемой с ней группой анцестрального таксона, можно говорить о полифилии. Если же сходство между группами дочернего таксона выражено слабее, а сходство между собой систематических групп исходного таксона выше, можно думать о монофилии. Третий случай, когда по исследуемым признакам сходство дочерних таксонов равно сходству между анцестральными таксонами, т. е. случай параллелизма, по сути дела, и является основой представлений об ограниченной полифилии (ср. Татаринов, 1976), тогда как принцип строгой монофилии приложим к ним с трудом (Татаринов, 1977).
При рассмотрении возможностей освоения новой адаптивной зоны двумя или большим числом систематических групп исходного таксона, занимающего соседнюю адаптивную зону, необходимо учитывать следующие параметры: степень таксономической и экологической близости этих групп, синхронность или асинхронность освоения ими новой зоны и территорию, на которой происходит этот процесс.
Если осваивающие новую зону группировки достаточно различны, т. е. далеки друг от друга по систематическому положению, то вопрос о полифилии или монофилии дочерних таксонов, по всей вероятности, не возникает. Различная организация предков будет определять и разные пути адаптации в новой зоне. Так, пути приспособления к вторичноводному образу жизни у представителей отряда Anura - Pipidae и представителей отряда Urodela - Proteidae части Ambystomidae, Amphiumidae, Cryptobranchidae и др. совершенно различны. У Urodela это приспособление всегда происходит посредством более или менее глубокой неотении, тогда как у Anura метаморфоз сохраняется и сохраняются свойственные этим отрядам различия организации, отчасти они даже усиливаются.
На уровне семейств и подотрядов организационные различия менее резко выражены, и пути адаптации к сходным условиям среды более близки. Так, приспособления к удлинению языка при выбрасывании, т. е. к дистантному захватыванию пищи у Bufonidae (подотряда Prococla) и Ranidae (подотряд Diplasiococla), происходят по-разному. У Bufonidae прогрессивно развиваются sinus sublingualis и тело языка (см. рис. 23, 24, 25), а у Ranidae - дивертикулы этой полости и рога языка (см. рис. 15, 26, 28) (Северцов, 1974). Различно формируется и взаимодействие мышцы - протроктора языка (m. genioglossus) с s. sublingualis. И у Bufonidae и у Ranidae эта мышца дифференцируется на мощную pars basalis, обеспечивающую напряжение и приподнимание прикрепленной к симфизу нижней челюсти проксимальной части языка и расходящуюся множеством пучков pars dorsalis, которая, опираясь на напряженную s. sublingualis, распрямляется и выбрасывает язык (см. рис. 16, 18, 23-28). У Bufonidae, кроме того, возникает еще одна порция - pars lateralis, подтягивающая основание языка к симфизу нижней челюсти (см. рис. 24). Однако формирование pars basalis у Ranidae и Bufonidae происходило независимо. Разными оказались морфология m. genioglossus и s. sublingualis и механизм их действия. У лягушек волокна базальной порции прикрепляются на дорсальной (передней) стенке лимфатической полости (см. рис. 16, 27). Ее сокращение растягивает эту лимфатическую полость и приводит к всасыванию в нее дополнительного объема лимфы из s. basihyoideus (см. рис. 16, 28). Повышение давления в s. sublingualis, обусловливающее ригидность языка и приводящее к его распрямлению под действием дорсальной порции m. genioglossus, создается исключительно сокращением мышц дна ротоглоточной полости (см. рис. 18), оказывающих давление на вентральную стенку s. sublingualis. У жаб волокна базальной порции прикрепляются к спинке языка, и ее сокращение сжимает s. sublingualis, дополняя действие мышц дна ротоглоточной полости. Дополнительное количество лимфы в полость не всасывается, а действие на нее pars dorsalis m. genioglossus диаметрально противоположно наблюдаемому у Ranidae.
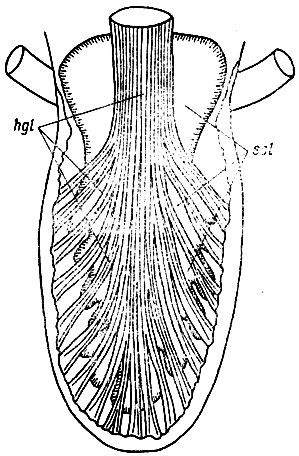
Рис. 23. Язык Bufo bufo с вентральной стороны. Строение m. hyoglosus (hgl) и лимфатической полости sinus sublingualis (ssl), инъецированной по Тейхману. Видно, что рога языка не выражены, а единая лимфатическая полость с короткими и широкими дивертикулами занимает практически всю площадь тела языка
Таким образом, у жаб и у лягушек сходная дифференциация мышц приводит к одинаковому результату - совершенствованию выбрасывания языка. Однако механизм действия мышц у обоих семейств различен, и один из другого не выводится. Даже если бы не было известно, что у Hylidae, форм не менее сухопутных и не менее продвинутых, чем два обсуждаемых семейства, дифференциация m. genioglossus на порции отсутствует, сами описанные различия позволили бы считать эволюцию языка Ranidae и Bufonidae параллелизмом и возникновение этой структуры дифилетическим.
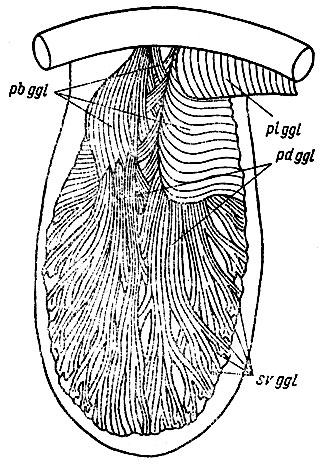
Рис. 24. Строение m. genioglossus у Bufo bufo. pbggl - pars basalis m. genioglossus - напрягает и приподнимает основание языка, одновременно надавливая на основание s. sublingualis и повышая в ней давление лимфатической жидкости, pdggl - pars dorsalis m. genioglossus. Пучки этой порции огибают напряженную лимфатическую полость, как канаты шкив блока и, сокращаясь, выпрямляют язык, поворачивают его концом вперед и выбрасывают, suggl - вентральный слой m. genioglossus (волокна обрезаны, полностью изображен на рис. 25). Этот слой, сжимая язык в поперечном направлении, проталкивает лимфатическую жидкость в s. sublingualis в свободный конец языка, обусловливая тем самым его удлинение, plggl - pars lateralis m. genioglossus, напрягая дно ротоглоточной полости смещает основание языка вперед к симфизу нижней челюсти, способствуя его выбрасыванию
Аналогично интенсификация дыхательных движений подъязычного аппарата приводит к разным, хотя и сходным, способам совершенствования его движений. Аспирация - втягивание воздуха в ротоглоточную полость достигается у Anura посредством поворота подъязычного аппарата передним краем вниз. Это движение происходит под действием m. sternohyoideus, оттягивающего подъязычный аппарат назад, и m. m. petrohyoidei, удерживающих на месте загнутые вверх задние рога подъязычного скелета (рис. 29). Однако поскольку указанные мышцы друг другу не параллельны, при их сокращении возникает сила, смещающая подъязычный аппарат дорсально, т. е. снижающая эффективность аспирации. Поэтому у более продвинутых семейств вырабатываются адаптации, предотвращающие это смещение. Последнее достигается путем приближения к параллельному расположению m. petrohyoideus III и т. sternohyoideus и возникновению механизма, фиксирующего ось вращения подъязычного аппарата. Первый эффект достигается посредством более дорсального расположения дистальных концов задних рогов, а следовательно, и места прикрепления m. petrohyoideus III. Однако у трех наиболее сухопутных из изученных семейств Anuna - Hylidae, Ranidae, Bufonidae это происходит по-разному. У Hylidae задние рога удлиняются без изменения угла их наклона относительно гиоидной пластинки. У Ranidae и Bufonidae расположение задних рогов становится почти вертикальным, но у Ranidae это происходит за счет изгиба вверх заднего края пластинки, а у Bufonidae - самих рогов (см. рис. 29).
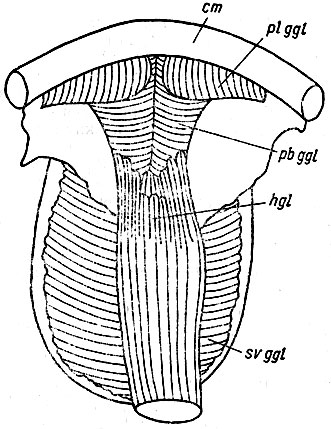
Рис. 25. Язык Bufo bufo с вентральной стороны: cm - нижняя челюсть. Обозначения, как на рис. 23 и 24. Видно расположение вентрального поперечного слоя m. genioglossus (svggl)
Фиксация оси вращения подъязычного аппарата достигается благодаря тому, что дорсальная сторона волокон m. sternohyoideus отщепляется от переднего края этой мышцы и прикрепляется у основания задних рогов, на уровне прикрепления m. petrohyoideus I. У Rana, но не у Hyla и Bufo эти волокна оформляются в самостоятельную головку. В то же время m. petrohyoideus II у Rana и Hyla становится слабее, а у Bufo исчезает. В результате m. petrohyoideus III и вентральная часть m. sternohyoideus создают момент вращения подъязычного аппарата, а прикрепляющиеся друг против друга m. petrohyoideus I и дорсальные волокна т. sternohyoideus фиксируют ось вращения (см. рис. 29).
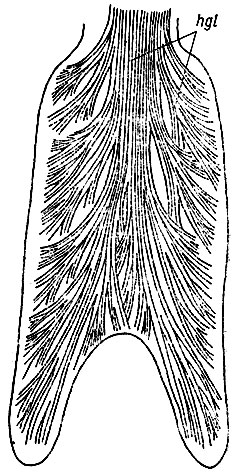
Рис. 26. Язык Rana temporaria с вентральной стороны. Строение m. hyoglossus (hgl). Видно расположение рогов языка. Поперечный слой т. genioglossus удален
Нетрудно заметить, что, как и при совершенствовании выбрасывания языка, один и тот же результат - повышение эффективности аспирации - достигается несколькими путями, что обусловлено не различиями направлений адаптации - это направление едино, а разной конструктивной основой, на которой параллельно формировался одинаковый результат. Иными словами, на уровне семейств отличить полифилию, доказав дивергенцию, при достаточной изученности материала не составляет особого труда. Естественно, на палеонтологическом материале, тем более фрагментарном и не допускающем функционального анализа, это сделать труднее. Однако, видимо, в любом случае своеобразие путей морфологических преобразований будет однозначно указывать на самостоятельность филогенеза групп. Даже на уровне таксонов относительно низкого ранга можно судить о том, возникло ли сходство полифилетически или является наследством от общих предков. Вышеприведенные примеры показывают, что к одним и тем же условиям среды могут приспособиться (более или менее одновременно) виды, принадлежащие к разным таксонам надвидового ранга.
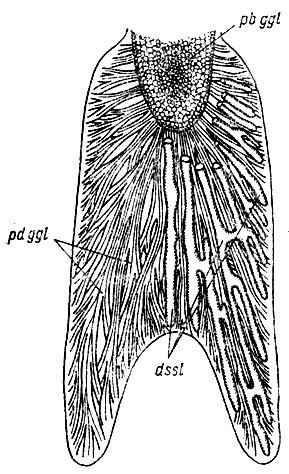
Рис. 27. Язык Rana temporaria с вентральной стороны Поперечный слой m. genioglosus и т. hyoglossus удалены. Видно расположение инъецированных по Тейхману дивертикулов s. sublingualis (dssl), пронизывающих толщу языка и его рогов между пучками дорсальной порции m. genioglossus (pdggl) и пучками п. hyogiossus, изображенным на рис. 26. Волокна базальной порции т. genioglossus (pbggl) оканчиваются на передней стенке sinus sublingualis (см. рис. 28). Их сокращение приводит к приподниманию базальной части языка и расширению лимфатической полости, в которую всасывается лимфа из s. basihyoideus (рис. 18, 28). Увеличение объема лимфы при давлении на нее основания мышц дна ротоглоточной полости приводит к повышению давления в s. sublingualis. Тем самым создается опора для досальной порции m. genioglossus, распрямляющей язык, как у Bufo bufo. Пучки дорсальной порции опираются на лимфатическую полость, как на шкив блока, и поворачивают дистальную часть языка, распрямляя его. Одновременно вентральный поперечный слой m. genioglossus сжимает язык в поперечном направлении, проталкивая лимфу по дивертикулам в его дистальную часть, что приводит к удлинению языка и его рогов
Вместе с тем несомненно, что по мере понижения таксономического ранга групп, эволюционирующих в одном направлении, понижается и возможность различения путей их адаптации к новым условиям среды. Иными словами, тенденция к монофилетической интерпретации происхождения надвидовых таксонов, даже если эта интерпретация не верна, подтверждается на рецентном материале. В противовес такому подходу можно выдвинуть дарвиновское представление о том, что чем ближе между собой формы, осваивающие одну и ту же среду обитания или сходные среды, тем интенсивнее между ними конкуренция. Конкуренция в свою очередь будет приводить к дивергенции этих групп, определяя увеличение различий между ними. Подобный процесс должен происходить, однако, при более или менее высокой насыщенности адаптивной зоны, в которую происходит вселение, и при симпатрическом ее освоении. При асинхронном освоении новой зоны может возникнуть ситуация асинхронного параллелизма, и группа, начавшая адаптироваться в новой зоне, либо "не пустит" другую группу в эту зону, либо, напротив, группа начавшая адаптироваться позже, вытеснит более ранних вселенцев. В обоих случаях вопрос о полифилии или монофилии не возникает, так как из двух групп остается и даст веер радиации в новой зоне лишь одна.
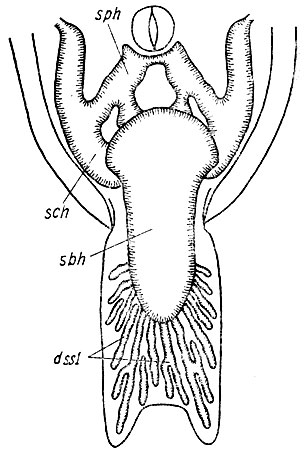
Рис. 28. Поверхностные лимфатические полости дна ротоглоточной полости Rana temporaria Видно расположение инъецированных по Тейхману дивертикулов s. sublingualis (dssl), сама полость полностью закрыта s. basihyoideus (sbh). Sch - s. ceratohyoideus, sph - sinus parahyoideus
Вероятность полифилии увеличивается, если предположить более или менее одновременное вселение в новую зону относительно близких групп, происходящее на разных территориях. Тот же эффект будет наблюдаться при синхронном и симпатрическом освоении разными таксонами субзон одной адаптивной зоны. Такая ситуация, отраженная в вышеприведенном примере с совершенствованием механизма дыхания Ranidae, Hylidae и Bufonidae, по-видимому, достаточно распространена: разные, но близкие по происхождению, а потому сходные группы будут осваивать сходные условия среды несколько различными путями, не конкурируя или слабо конкурируя между собой. Конкуренция будет возрастать лишь в процессе дивергенции их потомков уже в новой зоне. Различия исходной организации дают возможность каждой из группировок адаптироваться в новых условиях, не встречая конкуренции со стороны экологически близких вселенцев. Затем, уже по мере адаптивной радиации каждой из групп в новой зоне и их расселения, эти дочерние группы будут взаимодействовать между собой, расходясь по разным подзонам или вытесняя друг друга, но обе группы будут сосуществовать в новой и общей для них зоне. Наиболее ярким примером является сосуществование Reptilia и Раrаreptilia (Ивахненко, 1983; Михайлов, 1988). Подобный процесс возможен лишь при ненасыщенности зон, но эта ненасыщенность, видимо, должна быть характерным свойством любых вновь осваиваемых сред обитания. А. П. Расницын (1987) подчеркнул, что увеличение таксономического разнообразия связано с ослаблением конкуренции. Примером сосуществования разных по происхождению, но близких форм на одной территории может служить формирование фаун муравьев пустынь (Длусский, 1981). На каждом континенте пустынные фауны муравьев формировались из местных элементов, причем при освоении разных трофических ниш конвергентно возникал в разных группах (подсемействах и трибах) характерный облик тех или иных экологических форм.
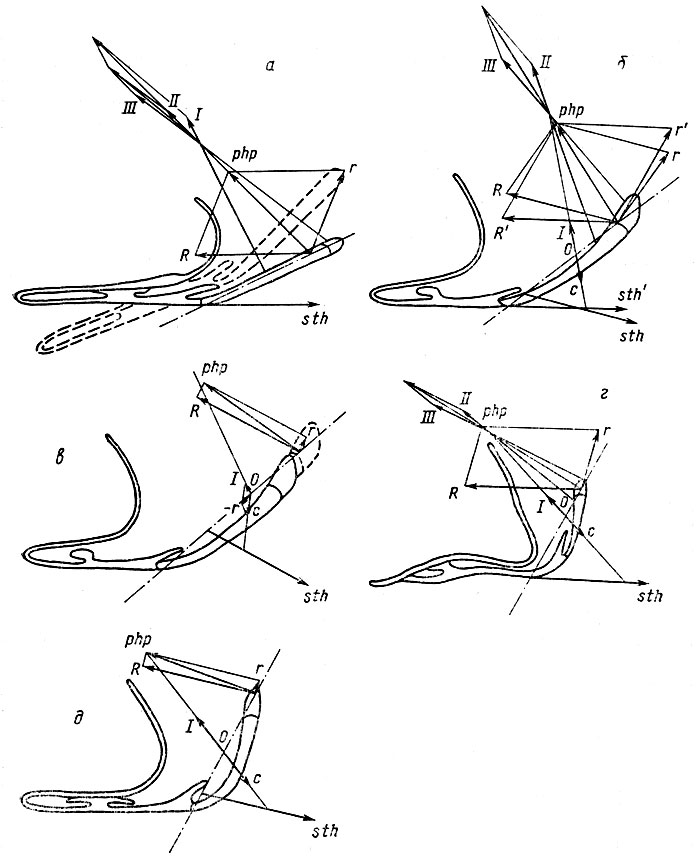
Рис. 29. Биомеханика опускания дна ротоглоточной полости Anura при аспирации. Подъязычный скелет (вид сбоку) и векторы участвующих в движении мышц у Bombina bombina (a), Hyla arborea (б, в), Rana temporaria (г) и Bufo calamita (d): sth - вектор m. sternohyoideus, ему противодействует вектор R, возникающий при разложении равнодействующей (php) m. m. petrohyoidei posteriores (I, II, III). Антипараллельные векторы sth и R обусловливают поворот подъязычного аппарата передним концом вниз (пунктир). Второй составляющей php является вектор г, снижающий эффективность аспирации: этот вектор приподнимает подъязычный аппарат, уменьшая тем самым амплитуду опускания дна ротоглоточной полости. При разрастании места прикрепления m. sternohyoideus назад и вверх по заднему рогу (рис. б) направление вектора этой мышцы меняется (sth' - sth), что приводит к уменьшению вектора r (r' - r). При более дорсальном расположении точки приложения php (рис. в) вектор г также уменьшается. Изменение точки приложения sth относительно точки приложения может достигаться либо за счет удлинения задних рогов, как у Hyla, либо за счет уменьшения угла их наклона относительно гипобранхиальной пластинки, как у Rana и Bufo. Обращает внимание, что более вертикальное положение задних рогов достигается у Rana изгибом гипобранхиальной пластинки, а у Bufo - самих рогов. По мере дорсального смещения вызывающий его вектор r быстро уменьшается (сравни б и в). Одновременно возникает и увеличивается противодействующий дорсальному смещению вектор r. Этот вектор, свойственный высшим Anura, обусловлен тем, что дорсальная часть m. sternohyoideus прикрепляется на заднем роге подъязычного скелета на одном уровне с m. petrohyoideus post I. При сокращении этих мышц (векторы с и I) возникает фиксированная ось вращения (О) подъязычного аппарата. Изменение положения этой оси под действием r и приводит к возникновению - r, предотвращающего дорсальное смещение и повышающего эффективность аспирации
Так, карпофаги представлены на разных континентах следующими формами
Неоарктика: Myrmicinae: Pogonomyrmex
Pheidolini: Veromessor, Novomessor
Solenopsidini: Solenopsis
Неотропика: Myrmicinae: Pogonomyrmex
Pheidolini: Pheidole, Elasmopheidole
Solenopsidini: Solenopsis
Палеарктика: Pheidolini: Messor, Pheidole, Oxyopomyrmex
Monomoriini: Monomorium
Tetramoriini: Tetramorium
Афротропика: Monomoriini: Monomorium
Tetramoriini: Tetramorium
Австралия: Monomoriini: Chelaner
Tetramoriini: Tetramorium
Pheidolini: Pheidole
Для этих форм характерны черты конвергентного сходства: мощные тупые жвалы, связанное с развитием жевательных мышц развитие затылочных углов головы; наличие псаммофора; вариабельность рабочей касты, часто доходящая до четкого диморфизма. Аналогичная система видов приведена для жизненных форм дневных зоонекрофагов и ночных зоофагов. Перечисленные роды карпофагов имеют различную распространенность. При анализе распространения видов Г. М. Длусский показал, что во всех пустынях фауна коадаптивных комплексов, включающая все три типа жизненных форм, складывалась автохтонно из местных семиаридных ценофобных видов, но затем в нее включались мигранты из других аридных зон, что приводило к усложнению комплексов. Иными словами, происходило смешение фаун, возникших более или менее синхронно, но аллопатрично. Таким образом, обсуждая первый из сформулированных выше вариантов, не удается выявить какие-либо запреты эколого-эволюционного характера, которые не позволяли бы считать возможным освоение новой адаптивной зоны двумя или несколькими более или менее близкими формами. Напротив, в литературе, касающейся рецентных форм, не удалось обнаружить указаний, подтверждающих строгую монофилию.
Отвечая на второй вопрос, необходимо подчеркнуть, что во всех приведенных примерах не возникает вопроса о таксономической принадлежности упомянутых форм. Даже в случае пустынных карпофагов подсемейства Myrmeciane никому не приходит в голову считать всех представителей этой эколого-морфологической группы единым таксоном. Видимо, подобные мнения могут возникать лишь в тех случаях, когда материал беден, а период освоения новой адаптивной зоны отдален во времени, как это часто бывает в палеонтологии. При неполноте материала возможны два альтернативных варианта: либо потомки групп будут восприниматься исследователем как единый таксон, и он будет настаивать на монофилетическом его происхождении; либо будут получены данные, позволяющие установить филогенетические связи компонентов этой группы с различными, параллельно эволюционирующими предками. В обоих случаях возникает, как отмечено выше, таксономическая, но не филогенетическая проблема, сформулированная в третьем вопросе.
В рамках дарвинистического подхода естественная система - это система "по происхождению", основанная на строгом соблюдении принципа генеалогического родства. Иными словами, требование строгой монофилии, согласно которому весь дочерний таксон берет начало от одного исходного вида, - это требование не теории филогенеза, а теории систематики. Поэтому доказательство полифелитического происхождения того или иного таксона ставит вопрос об его интерпретации. В систематике имеются прецеденты разделения подобных таксонов. Так произошло с Rodentia и Lagomorpha, Reptilia и Parareptilia и рядом других групп. Однако подобная процедура, по-видимому, всегда будет встречать сопротивление, так как любая крупная перестройка сложившейся системы крайне затрудняет пользование ею. "Умеренно-консервативная позиция", предлагаемая для подобных ситуаций Л. П. Татариновым (1976, с. 206), вероятно, наиболее приемлема. Формулирование тех или иных принципов систематики в связи с полифилетическими таксонами не входит в задачу данной книги, посвященной направленности эволюции. Поэтому, видимо, целесообразно лишь подчеркнуть еще раз, что, насколько об этом можно судить, не существует запретов для перехода в новую адаптивную зону нескольких достаточно близких (экологически и филогенетически) групп исходного таксона. Но сама по себе проблема полифилии или монофилии - это проблема не теории филогенеза, а теории систематики.
Выше рассмотрен круг вопросов, связанный с традиционными подходами к анализу проблемы монофилии и полифилии. Однако в последние два десятилетия эта проблема получает несколько иное звучание в связи с достижениями молекулярной биологии. Открытие явления латерального переноса генетического материала привело к еще одной ее трактовке. Уже не гибридизация надвидовых таксонов, а перенос между ними отдельных генов обсуждается как проблема полифилетичности генома. Указанный аргумент в пользу полифилии обсуждать, по-видимому, преждевременно. Распространенность трансдукции в естественных условиях даже среди прокариот не выяснена. Тем более неизвестно, насколько привнесение отдельных генов в геном единичной особи какого-либо другого вида будет влиять на дальнейшую эволюцию этого вида. По сути дела, привнесение нового гена в геном равносильно единичной мутации и не вносит ничего нового в существующие представления об эволюционном процессе. Поэтому применение термина "полифилия" к формам, возникшим на основе трансвидового переноса генов, даже если такие формы будут обнаружены в природе, всегда будет чисто формальным. Сказанное, естественно, не умаляет огромного значения искусственного введения генов в чужеродный геном - "генной инженерии", имеющей решающее значение в биотехнологии, очень важной для изучения биосинтеза и ряда других проблем молекулярной биологии. Однако с точки зрения эволюции - это всего лишь один из путей возникновения генетической изменчивости.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'