2.6.1. Специализация
2.6.1.1. Теломорфоз
Этим термином И. И. Шмальгаузен (1969) обозначил специализацию в том смысле, в каком ее обычно понимают экологи: специализацию по питанию (олиго- и монофаги) и по местообитаниям (стенобионтные формы). Теломорфная, как и любая другая специализация, по И. И. Шмальгаузену (1982), представляет собой приспособление в более узких (по сравнению с предками) условиях окружающей среды, т. е. одностороннее приспособление. В качестве примеров теломорфной специализации И. И. Шмальгаузен приводил круглоротых с их своеобразными адаптациями к полухищному, полупаразитическому способу питания; ленивцев и муравьедов из млекопитающих к т. п. Ранее было отмечено (см. гл. 2.5), что специализация как процесс представляет собой сужение адаптивной зоны, т. е. более полное приспособление к узкому диапазону внешних условий.
Этот путь адаптации дает группе, идущей по пути теломорфоза, выигрыш в конкуренции с соседними таксонами, так как, с одной стороны, снижает конкуренцию между ними, а с другой - повышает конкурентоспособность "специалистов" в свойственной им узкой зоне. В этом, собственно, и заключается биологическая прогрессивность, позволяющая специализированным формам нара-щивать численность (биомассу) по мере снижения конкуренции с соседними таксонами и совершенствования механизмов использо-вания той среды, к которой они адаптируются.
Противодействовать биологическому прогрессу специализированных форм, или, точнее, сдерживать его, будут главным образом две группы факторов: ширина самой адаптивной зоны, т. е. ограниченность ресурсов, доступных стенобионтным формам, и коадаптивная с ними эволюция групп, как занимающих соседние адаптивные зоны, так и являющихся компонентами их собственной зоны. Поэтому биологический прогресс для специализированных форм - понятие часто весьма относительное. Для многих теломорфных (и не только теломорфных - см. ниже) таксонов он означает не более чем возможность выжить, в то время как менее специализированные родственные формы вымирают. Примером подобной специализации среди Anura могут служить Pipidae. Переход этого семейства к вторично-водному образу жизни, несомненно, представляет собой сужение адаптивной зоны предков. В процессе специализации Pipidae утратили способность существовать на суше, а потому - значительную часть своих экологических возможностей и эволюционных потенций. Специализация этих животных сопровождалась глубокой перестройкой организации. Pipidae утратили язык, что обусловило резкое таксономическое противопоставление этого семейства всем остальным Anura, подразделение последних на Aglossa (Pipidae) и Phaneroglossa (все остальные бесхвостые амфибии). Компенсаторно у Pipidae развился очень своеобразный способ питания. Передними конечностями африканские представители этого семейства разрывают и взбаламучивают ил и выхватывают оттуда пищу - мелких беспозвоночных, зажимая их между двумя соседними пальцами. Схваченная таким образом пища отправляется лапами в рот. Всасывание добычи с током воды, свойственное при питании под водой амфибиям и унаследованное ими от кистеперых рыб (Северцов, 19806), у родов Xenopus и Hymenochirus практически не наблюдается. У Pipa этот способ питания имеется, но механизм его иной, чем у Phaneroglossa. Если учесть, что использование для питания пальцев передних конечностей свойственно среди позвоночных млекопитающим и птицам, то своеобразие специализации Pipidae будет еще более наглядным.
В то же время сохранение свойственного амфибиотическим Anura механизма вентиляции легких в условиях характерного для Pipidae длительного пребывания под водой обусловило прогрессивное развитие и легких, и подъязычного аппарата. У Pipidae впервые в ряду позвоночных возникает сложноячеистая структура легких, с мелкими полостями, сходными с альвеолами, и охрящевение стенок канала, соединяющего легкие с гортанью, что делает его похожим на трахею (De Jong, 1972). Оба признака у других амфибий не встречаются. Подъязычный хрящ достигает колоссальных размеров, вдаваясь назад, далеко в глубь полости тела (см. рис. 19), что позволяет одним движением нагнетательного насоса наполнять легкие. В связи с дальнейшим по сравнению с амфибиотическими Anura приспособлением к плаванию изменены строение таза и задних конечностей.
Наконец, у Pipa americana сформировался широко описанный в литературе, крайне своеобразный механизм заботы о потомстве. Оплодотворенные икринки имплантируются в кожу спины: кожа набухает, образуя камеры, каждая из которых заключает одну икринку. Онтогенез потомства, вплоть до окончания метаморфоза, протекает в этих камерах, откуда выходят вполне сформировавшиеся сеголетки.
Из приведенного краткого описания теломорфных преобразований организации Pipidae видно, что часть из них представляет резко "продвинутые" признаки, т. е. признаки, имеющие аналогии у высших по отношению к амфибиям форм. При желании эти признаки можно было бы рассматривать как филогенетические предварения, а самих Pipidae (не учитывая их экологии) как "пророческие формы". Беда только в том, что эти филогенетические предварения служат адаптациями к жизни в воде, хотя "предваряют" признаки сухопутной адаптации высших животных.
Другая часть признаков представляет собой своеобразные уклонения от типичной для Anura морфологии, и, наконец, третий комплекс признаков - это признаки примитивности, к каковым, например, относятся амфицельные позвонки, примитивное строение крестца и ряд других. В целом же морфология Pipidae характеризует, с одной стороны, аберрантность этого семейства, а с другой - мозаичность организации входящих в него форм. Аберрантность, своеобразие морфологии, по-видимому, наиболее яркая характеристика специализации, как сужения адаптивной зоны. Приспособление к узким, а потому своеобразным условиям среды диктует и своеобразие организации. В то же время необходимо учитывать, что сужение адаптивной зоны одновременно является, как правило, и упрощением среды обитания. Действительно, в обсуждаемом примере с Pipidae одностороннее приспособление к жизни в воде делает ненужным весь комплекс адаптаций к жизни на суше, с чем, в частности, связана редукция языка. В наибольшей степени упрощение организации свойственно катаморфозу (см. ниже), но и при теломорфозе оно присутствует, хотя обычно менее ярко выражено.
Как отмечено выше, Pipidae свойственна довольно заметная гетеробатмия организации, являющаяся для этих животных выражением их примитивности. Считается, что Pipidae - одна из наиболее рано отделившихся ветвей веера адаптивной радиации Anura (Duellman, 1975; Noble, 1931). В соответствии с представлениями A. Л. Тахтаджана (1966) подобная гетеробатмия по мере филогенеза таксона должна постепенно утрачиваться благодаря формированию системы координаций. По-видимому, можно предполагать два различных филогенетических механизма формирования теломорфной специализации. При первом из них в процессе адаптивной радиации группы, осваивающей новую адаптивную зону, часть ветвей этой радиации попадает в краевые, узкие, аберрантные (экологически) подзоны и вынуждена в них адаптироваться, т. е. специализироваться, так как продолжающаяся конкуренция с другими ветвями того же веера адаптивной радиации или других таксонов не позволяет им расширить или сменить ту узкую подзону, в которую их завела конкуренция с соседними систематическими группами.
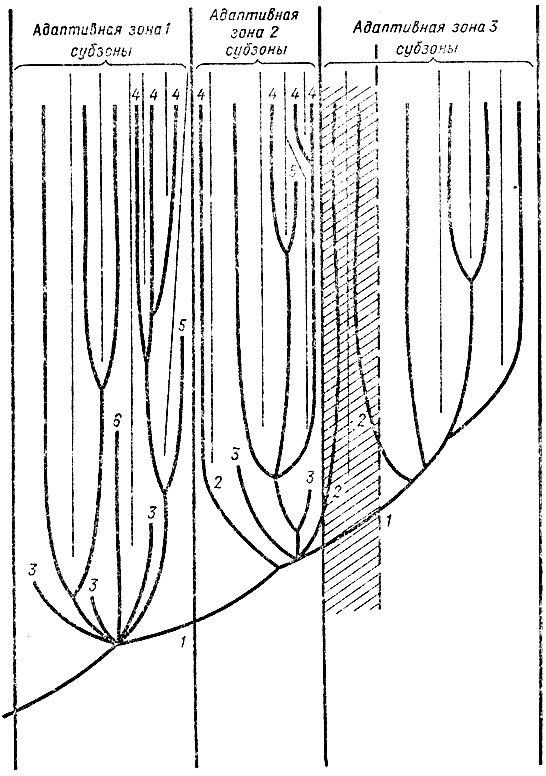
Рис. 30. Схематическое изображение возможных вариантов филогенеза надвидовых таксонов Три соседние адаптивные зоны последовательно осваиваются в ходе прогрессивной эволюции. Освоение 2-й происходит посредством смены зон (алломорфоз), освоение 3-й - посредством расширения (ароморфоз). Часть зоны анцестрального таксона, ставшая частью зоны дочернего таксона, заштрихована. Расширение зоны не препятствует существованию в унаследованной от предков ее части филумов анцестрального таксона (напр. двоякодышащие и амфибии). 1 - ранняя специализация, 2 - ранняя специализация, переходящая в позднюю специализацию, 3 - филумы, вымершие в результате ранней специализации и (или) в результате вытеснения промежуточных форм, 4 - поздняя специализация, 5 - специализированные филумы, вымершие в результате изменения среды, 6 - филум, вымерший в результате вытеснения его более приспособленным, параллельно эволюционирующим таксоном (инадаптивная эволюция)
Подобную специализацию можно назвать ранней (рис. 30; гл. 2.5). Для нее характерны высокая степень гетеробатмии и сохранение черт примитивности, т. е. несформированность системы координаций. Следовательно, ранней специализации свойственно сохранение, несмотря на специализированность, эволюционной пластичности, прежде всего за счет несформированности координаций.
Поздней специализацией можно назвать тот случай, когда специализированные формы возникают в процессе дальнейшего дробления субзон дочерних таксонов, в то время как освоение адаптивной зоны данной систематической группой уже закончено. Подобная специализация, происходящая на фоне уже сформированной или в основном сформированной системы координаций, характеризуется гораздо более низкой гетеробатмией, а потому и снижением эволюционной пластичности. Примерами поздней специализации могут служить роющие млекопитающие: настоящие кроты из насекомоядных, цокоры, слепушонки и слепыши из грызунов. Эти примеры приводил для характеристики теломорфоза И. И. Шмальгаузен (1982). Для всех перечисленных форм характерно мощное развитие адаптаций к роющему образу жизни (см. Гамбарян, 1960), но не удается выделить признаков гетеробатмии.
Как сказано ранее (см. гл. 2.5), чем выше эволюционная пластичность группы, т. е. чем в большем числе направлений она может эволюционировать, тем менее вероятно ее вымирание. Эволюционная пластичность складывается из двух компонент: степени мультифункциональности органов, определяющих связи с окружающей средой (см. гл. 2.4), и степени координированности организации в неиерархических координационных цепях, сокращающей число возможных направлений перестройки этих цепей (см. гл. 2.4). Снижение мультифункциональности характерно и для ранней, и для поздней специализации, но сформированность системы координаций - свойство в первую очередь поздней специализации.
Как показал И. И. Шмальгаузен (1982), для специализации вообще и для теломорфоза в частности характерно постепенное замедление темпов эволюции, которое он объяснял усилением стабилизирующего отбора с продвижением таксона по пути специализации. Начальные этапы специализации, особенно ранней, должны, по-видимому, протекать относительно быстро. По мере освоения новой зоны и ее сужения происходит снижение мультифункциональности тех органов, которые определяют адаптацию к новым условиям. Вместе с тем достигается все более полное и совершенное приспособление специализирующихся форм к более узкой адаптивной зоне. В результате снижается интенсивность конкуренции с группами, занимающими соседние адаптивные зоны. В снижении этой конкуренции и состоит, по-видимому, один из важнейших адаптивных эффектов специализации и его причина. Чем выше эврибионтность видов (или надвидовых таксонов), находящихся на одном трофическом уровне, тем по большему числу параметров среды они между собой конкурируют. Формирование стенобионтности специализирующихся форм приводит к снижению конкуренции за одни и те же условия среды, так как таких условий становится меньше.
Вместе с тем продолжающаяся адаптация к условиям собственной узкой зоны приводит к повышению численности и интенсификации индивидуальной конкуренции. Поэтому особи, отклоняющиеся от уже сформировавшегося пути специализации, оказываются в худших по сравнению с нормой условиях борьбы за существование и чаще элиминируются. Действует строгий движущий отбор, совершенствующий признаки специализации. В этом смысле справедливо правило специализации Ш. Депере (1915), согласно которому группа, вставшая на путь специализации, может эволюционировать только в сторону дальнейшей специализации. Однако по мере совершенствования специализированных признаков интенсивность отбора постепенно снижается, так как дальнейшее развитие этих признаков уже не дает выигрыша в борьбе за существование. В результате возрастает роль стабилизирующего отбора, который, с одной стороны, через перестройку морфогенетических корреляций повышает целостность организма (Шмальгаузен, 1969, 1982), что выражается в формировании системы координаций, а с другой - останавливает эволюцию высоко специализированных форм (нормализующий эффект). Персистентные формы - это, как правило, высоко специализированные формы.
Разница между ранней и поздней (см. рис. 30) специализацией по мере формирования системы координаций, по-видимому, теряется. Группа, достигшая высокой степени специализации, будет существовать в своей узкой адаптивной зоне до тех пор, пока существует эта зона. Иными словами, специализация не обязательно ведет к вымиранию. Индивидуальная изменчивость высоко специализированных, в том числе и персистентных, форм ничуть не ниже, чем любых других. Это показано непосредственными исследованиями у Limulus и других видов (Левонтин, 1978). Приспособленность специализированных форм в свойственных им условиях среды очень высока. Однако поскольку далеко зашедшая специализация ведет к снижению эволюционной пластичности, изменение адаптивной зоны специализированных форм (и только оно) способно привести эти формы к вымиранию: они просто не успевают (не могут) перестроиться вместе с изменениями окружающей среды. Отсутствие соответствующих функций приведет к тому, что отбор по признаку, даже при изменении внешних условий, происходить не будет, а ограничения, накладываемые координациями, еще более снизят возможность перестройки организации специализированных форм. Специализированные теломорфные формы в рамках таксона данного ранга, так же как и в рамках конкретных фаун и флор, по-видимому, никогда не бывают многочисленными. Для ранней специализации это объясняется тем, что в аберрантные субзоны, именно в силу их аберрантности, а потому редкости, могут попасть лишь немногие виды исходного веера адаптивной радиации.
Легче представить себе постепенное дробление адаптивной зоны в ходе дальнейшей эволюции таксона и широкое распространение поздней специализации (см. рис. 30). Однако этому будут постоянно препятствовать: неравномерность процессов биологического прогресса (Гептнер, 1965) и процессы вытеснения инадаптивных форм более медленно эволюционирующими, а потому лучше приспособленными их конкурентами. Экологические и морфологические аспекты специализации, связанные с освоением новых адаптивных зон, были обсуждены в гл. 2.5, как и различия ранней и поздней специализации. В заключение следует лишь сказать, что теломорфоз представляет собой конгломерат очень разнообразных по своей экологической значимости и организационной обусловленности направлений адаптации, объединяемых лишь одним параметром - возрастанием стенобионтности по мере филогенеза.
2.8.1.2. Гиперморфоз
Под гиперморфозом И. И. Шмальгаузен (1982, 1969) понимал "переразвитие организма", относя к этому типу специализации не только гигантизм как таковой, но и переразвитие отдельных частей организма, например рогов у гигантского торфяного оленя - Megaloceros, клыков у саблезубых кошек и у бабируссы, пластин на спине стегозавров и т. п. И. И. Шмальгаузен подчеркивал, что подобное переразвитие представляет собой один из случаев инадаптивной эволюции, так как переразвитие частей организма приводит к дискоординации, т. е. снижению целостности организма. Причины такой дискоординации И. И. Шмальгаузен видел в быстрых темпах гиперморфной эволюции, благодаря чему не успевает перестроиться система онтогенетических корреляций, и в результате продления аллометрического роста за те пределы, в которых он происходил у предков, возникают диспропорции. Это характерно не только для переразвития отдельных признаков, но и для гигантизма как такового. И. И. Шмальгаузен подчеркивал, что для гигантских динозавров, как и для гигантских млекопитающих, по мере увеличения их размеров в ходе филогенеза относительные размеры головного мозга уменьшаются, что характерно для изменения соотношения массы этого органа и массы тела в онтогенезе всех позвоночных. В дис- координации И. И. Шмальгаузен видел одну из основных причин недолговечности гиперморфных филумов.
Поскольку гигантизм - гипертелия (Соре, 1904) - представляет собой, по-видимому, наиболее часто встречающийся случай гиперморфоза, необходимо, во-первых, выяснить, почему этот тип биологического прогресса является специализацией и, во-вторых, в каких условиях возникает это направление эволюции. И у животных, и у растений гигантизм может развиваться лишь при наличии кормовой базы (для растений - света, воды, минерального питания), достаточной для прокормления гигантских форм при сохранении плотности их населения, обеспечивающей нормальное размножение. Поэтому, если теломорфоз - специализация по способу добывания пищи, то гиперморфоз - специализация прежде всего по количеству пищи. Часть случаев переразвития отдельных органов, представляющих собой средства нападения и защиты (клыки саблезубых кошек, костные воротники цератопсов, пластины на спине стегозавров, рога титанотериев и т. п.), сближают гиперморфоз и теломорфоз. Возможно, все подобные признаки, указывающие на пищевую специализацию или, напротив, защиту от специализированных к добыванию крупной добычи (см. ниже) хищников, следовало бы отнести к теломорфозу. Во всяком случае не удается найти принципиальных различий между переразвитием клыков у саблезубых кошек - гиперморфоз, по Шмальгаузену (1982), и переразвитием резцов у слепышей или у цокоров - теломорфоз, по Шмальгаузену (1982), и переразвитием клыков у моржей, которое отчасти связано с добыванием пищи, отчасти (у самцов клыки крупнее) с половым диморфизмом.
И. И. Шмальгаузен проводил это разграничение на основе единства механизмов, приводящих к выработке подобных приспособлений, и гигантизма как такового. Для животных - это, по всей вероятности, коэволюция пары хищник - жертва. Если жертва изначально крупнее хищника, т. е. мало доступна ему, то хищник может переключиться на добывание молодых (более мелких) животных. В этом случае наиболее эффективный способ снижения возрастной элиминации жертвы - ускорение роста, что позволяет быстрее достигать размеров, защищающих от хищника. Тогда у хищника должен так же начинаться отбор на увеличение размеров, снова стимулирующий ускорение роста и увеличение размеров жертвы. Такой процесс будет продолжаться до тех пор, пока кормовая база жертвы и ограничения, накладываемые физикой или кинетикой, допускают увеличение ее размеров. Естественно, параллельно с увеличением размеров у жертвы развиваются средства защиты, а у хищника - средства нападения. Прекрасными иллюстрациями к такому направлению эволюции служат филогенетические ряды цератопсов, титанотериев и многие другие. Подобная сопряженная эволюция возможна только при отсутствии развитых механизмов группового поведения, т. е. когда хищник охотится в одиночку на одиночную жертву. При наличии групповой обороны, как это свойственно большинству современных копытных, и групповых способов охоты, например у волков и львов, отбор на увеличение размеров возникать не будет.
У растений причины гиперморфоза, естественно, отличаются от тех, которые ведут эволюцию в этом направлении у животных. Однако и у них, по-видимому, гиперморфоз все же связан с возрастной элиминацией. На это указывает то, что секвойевые леса представляют собой пирогенный климакс.
Другой класс явлений переразвития отдельных органов и отчасти, видимо, гигантизма представляют собой случаи переразвития вторично половых признаков, таких как рога Megaloceros, бивни мамонтов, завивавшиеся в спираль, клыки бабируссы и др. И. И. Шмальгаузен (1982) отмечал, что подобные признаки не дают их носителям выигрыша в борьбе за существование. Такая точка зрения объясняется тем, что в конце 30-х гг., когда И. И. Шмальгаузен писал "Пути и закономерности эволюции", внутрипопуляционные отношения были еще очень слабо изучены, что не позволяло в должной мере оценить их значение в эволюции, в том числе вторичнополовых признаков. Между тем для особей полигамных видов вторичнополовые признаки имеют не меньшее, если не большее значение, чем признаки, определяющие приспособленность в межвидовых отношениях. Поэтому переразвитие вторичнополовых признаков означает лишь возрастание роли внутривидовых отношений по сравнению с межвидовыми. Однако говорить в данном случае о специализации достаточно трудно. Едва ли можно считать, что пенженские популяции лося, обладающие наиболее сильно развитыми рогами, специализированы сильнее, чем любые другие популяции лосей палеарктики.
Кроме того, С. А. Северцов (1951) подчеркивал, что количественные характеристики вторичнополовых признаков, например величина рогов у Cervidae, обратимы. Так, при интенсивном выборочном отстреле наиболее болынерогих благородных оленей в странах Центральной Европы, что было связано с погоней за трофеями при спортивной охоте, начался отбор на "беднорогость", и олени того же возраста стали обладать рогами с меньшим количеством отростков. В настоящее время в ЧСФР путем выплаты премий за отстрел оленей и косуль с мелкими и уродливыми рогами ведется селекция на увеличение рогов. Если бы переразвитые вторичнополовые признаки, в частности рога Megaloceros, снижали относительную приспособленность старых самцов, обладавших наиболее крупными рогами, видимо, начался бы отбор против этого признака. Поэтому, во-первых, трудно предполагать, как это иногда делается в популярной литературе, что причиной вымирания гигантского торфяного оленя были его огромные рога, а во-вторых, возникает сомнение в том, насколько вообще переразвитие вторичнополовых признаков нарушало целостность организма и служило причиной вымирания видов, обладавших такими признаками.
Так или иначе гиперморфоз в том объеме, как его трактовал И. И. Шмальгаузен (1969, 1982), представляется явлением гетерогенным. Переразвитие отдельных органов, связанное с добыванием пищи, защитой от хищника или адаптацией к аберрантным местообитаниям, неотличимо от теломорфоза. Переразвитие вторично-половых признаков едва ли можно рассматривать как специализацию. Следовательно, гиперморфозом может считаться лишь гигантизм. Поэтому многие гиперморфные организмы сочетают в себе признаки гиперморфоза - гигантизма и теломорфоза.
Гигантизм действительно приводит к дезинтеграции организма, причем не к сохранению примитивной мозаичной организации, а именно к разрушению или искажению динамических или топографических координаций, что, возможно, снижает конкурентоспособность гиперморфных форм. Однако считать дискоординацию основной причиной вымирания едва ли возможно. Согласно представлениям А. Л. Тахтаджана (1966), по мере филогенеза гетеробатмии утрачивается (см. выше), т. е., по-видимому, не существует препятствий к тому, чтобы у гигантских форм складывались новые системы координаций. Как справедливо указывал И. И. Шмальгаузен (1982), гигантские формы обычно (в филогенетическом масштабе времени) недолговечны. Они быстро возникают и быстро вымирают. Однако показать, что гиперморфные группы инадаптивны и поэтому вытесняются более мелкими и более координированными формами, видимо, очень трудно.
По всей вероятности, гигантизм, как и теломорфная специализация, может оказаться причиной вымирания в силу экологических причин, т. е. соотношений организации и условий среды, а не особенностей организации. Действительно, количественные характеристики кормовой базы подвержены гораздо более сильным колебаниям, чем качественные, т. е. состав пищи. Поэтому кормовая база гиперморфных филумов должна быть существенно менее стабильной, чем теломорфных. Если теломорфоз при специализации по питанию ведет к формированию стабильной, независимой от конкурентов кормовой базы, то гиперморфоз не избавляет от пищевой конкуренции с более мелкими формами, тогда как зависимость гигантских форм от количества пищи усугубляется. Нестабильностью кормовой базы в сочетании с низкой плодовитостью и следует, видимо, объяснять недолговечность гиперморфных таксонов.
Одним из наиболее наглядных подтверждений вышесказанному служит вымирание пещерного медведя. Это гигантское животное было всеядным, на что указывает строение его зубов (Северцов, 1951). В целом этот мощный всеядный зверь, по-видимому, не имел врагов в современной ему фауне (может быть за исключением человека) и в период между миндельрисским и риссвюрмским оледенениями жил в условиях экологического оптимума. Об этом свидетельствуют сильная индивидуальная изменчивость остеологических остатков, обнаруженная О. Абелем (Abel, 1931) при раскопках в Драконовой пещере. Как показал И. И. Ежиков (1933), повышенная изменчивость является одной из характеристик экологического оптимума вида. Высокая изменчивость пещерных медведей свидетельствует, в частности, о том, что утрата эволюционной пластичности представляет собой не генетическую, как считали некоторые исследователи (Denepe, 1915; Соре, 1904; Doljo, 1893), а эколого-морфологическую характеристику таксона. Однако наиболее наглядным свидетельством того, что пещерный медведь жил в условиях экологического оптимума, является обнаружение О. Абелем (Abel, 1931) большого числа скелетов старых особей, умерших, по-видимому, от старости в своей пещере.
Причиной вымирания пещерного медведя, как показал С. А. Северцов (1951), послужило оскудение кормовой базы, произошедшее в связи с началом риссвюрмского похолодания. Более продолжительные зимы и весенняя бескормица оказались губительными прежде всего для медвежат. При низкой рождаемости - не более одного медвежонка, что устанавливается по скелетам плодов в неразродившихся самках, гибель молодняка привела к вымиранию вида в целом.
Наиболее крупные из современных млекопитающих - усатые киты, по-видимому, также могут служить доказательством, что кормовая база служит основным параметром, определяющим стабильность адаптивной зоны гиперморфных форм. Планктофагия в условиях Мирового океана оказалась очень стабильной кормовой базой. Лишь истребление человеком подорвало численность Mistacoceti. Напротив, судьба слонов, особенно африканских, не может рассматриваться в качестве модели гиперморфоза. Они подвергались прямому истреблению, а кроме того, сокращалась территория их обитания в результате окультуривания ландшафта. Поэтому выделить экологические причины их вымирания (сокращения численности) достаточно трудно.
Подводя итог вышесказанному, следует еще раз подчеркнуть, что гиперморфоз в том объеме, как сформулировал это направление эволюции И. И. Шмальгаузен (1982, 1969), представляет собой гетерогенное явление. Часть случаев гиперморфоза может быть отнесена к теломорфозам, часть (переразвитие вторично половых признаков) трудно считать специализацией как таковой, и лишь гигантизм оказывается действительно своеобразным типом специализации по количеству пищи. Вымирание гигантских форм обусловлено нестабильностью их кормовой базы, а не дискоординацией их организации. Дискоординация, видимо, с успехом компенсировалась снижением количества врагов, или, что то же самое, - повышением индивидуальной экологической устойчивости в условиях достаточной обеспеченности пищей (К-стратегия размножения, Пианка, 1981).
Если гигантизм как специализация возникает при направлении отбора, ведущем к укрупнению размеров в системе хищник - жертва, то при изначально обратном соотношении размеров - жертва мельче хищника - может возникнуть отбор на специализацию, противоположную гиперморфозу, - отбор на дальнейшее уменьшение размеров тела жертвы и соответственно хищника. В слабой форме это направление филогенеза можно проследить на копытных и тиграх индомалайского архипелага. В более яркой форме это прослеживается на насекомоядных (например, бурозубки-крошки), мышевидных грызунах (мыши-малютки) и хищниках из семейства куньих (ласка, в меньшей степени - горностай.)
Такому уменьшению размеров, которое по образцу номенклатуры И. И. Шмальгаузена можно было бы назвать наноморфозом, ставят предел физиологические ограничения: для гомойотермных животных - это соотношение объема и поверхности, неблагоприятное из-за высоких теплопотерь; для пойкилотермных рептилий - слишком быстрая потеря тепла и ограничение времени активности; для амфибий - высокая влагоотдача через кожу. Уменьшение размеров тела для гомойотермных животных с экологической точки зрения почти то же самое, что и гигантизм. Из-за больших потерь тепла подобные животные могут существовать только в условиях обильной кормовой базы, состоящей из доступных им по размерам объектов, или вырабатывать гетеротермию для перенесения неблагоприятных (малокормных) периодов, похолоданий и т. п. И для гомойотермных и для пойкилотермных животных ианоморфоз означает специализацию по местообитанию - выбор условий или микроусловий с оптимальными режимами температуры и влажности. Особенно характерна подобная специализация для беспозвоночных.
Однако у позвоночных (ноне Artropoda) наноморфоз - явление, по-видимому, еще более редкое, чем гиперморфоз. Дело в том, что для хищника уменьшение размеров тела обычно означает расширение кормовой базы - возможность использовать в пищу беспозвоночных. Поэтому по мере уменьшения размеров селективный пресс хищника ослабевает, а отсюда снижается стимул для дальнейшего уменьшения размеров жертвы. Истинная распространенность наноморфоза в филогенезе и животных и растений едва ли может быть сколь-нибудь надежно оценена. Останки мелких животных фоссилизируются лишь в редких случаях. Напротив, случаи гиперморфоза благодаря легкой фоссилизации костей крупных животных легко выявляются палеонтологами, и можно думать, что большинство случаев гигантизма уже известно, тогда как наноморфоз может быть показан только на рецентных формах.
2.6.1.3. Катаморфоз
Понятие катаморфоза, согласно И. И. Шмальгаузену (1982, 1969), совпадает с понятием общей дегенерации (Северцов, 1939, 1967). Этот тип специализации возникает при попадании того или иного филума в более простые условия среды, чем те, в которых обитали их предки. Обычно катаморфоз связан с переходом к сидячему или паразитическому образу жизни.
Более простые соотношения с внешней средой ведут к утрате большей части адаптаций, необходимых предкам, и к вторичному упрощению организации. При катаморфной эволюции наиболее четко прослеживается свойство специализации приводить к утрате ранее сформировавшихся, свойственных предкам ароморфозов. Это свойственно и поздней теломорфной специализации и особенно катаморфозу. Пользуясь общеизвестным примером катаморфоза - организацией асцидий (Северцов, 1939, 1967; Шмальгаузен, 1982, 1969), нетрудно показать, что при переходе к седентарному образу жизни эти животные утрачивают именно те признаки, которые позволяют на основе организации свободно плавающих личинок причислить их к хордовым. Утрачиваются хорда, метамерная мус-кулатура, в значительной мере нервная система и ряд других органов.
И. И. Шмальгаузен (1982) подчеркивал, что катаморфоз ведет к утрате и упрощению большей части биологических - динамических и топографических координаций и к снижению целостности организации. Последнее положение, видимо, не совсем точно. Утрата ставших ненужными в более простой, а потому и более стабильной среде органов, а отсюда и утрата координаций этих органов с другими частями организма действительно связана с глубокой перестройкой всех (или большей части) внутриорганизменных зависимостей. Однако это не означает дискоординацию оставшихся и прогрессивно развивающихся признаков. Напротив, упрощение координационных связей обеспечивает становление нового уровня целостности организма, что и наблюдается как у седентарных форм - тех же асцидий, у которых развитие фильтрационного аппарата глотки прекрасно коррелировано с развитием выводного сифона, присосок, кровоснабжением этих органов и т. п., так и у паразитических форм.
Возможно, однако, что эффект хорошей координированности организации катаморфных групп представляет собой следствие их длительной эволюции в свойственных им адаптивных зонах. Боль-шая часть древних групп, позволяющих в какой-то мере судить о соотношениях и происхождении типов животных, таких как Pterobranchia, Pogonophora, Ascidia, Cephalochordata и др., представляют собой катаморфные формы, дожившие до современности благодаря существованию в упрощенных стабильных условиях среды.
Вместе с тем необходимо отметить, что катаморфоз (общая дегенерация) связан с теломорфозом непрерывным спектром переходов. Уже для асцидий достаточно трудно отделить адаптации к упрощенной среде обитания, т. е. собственно катаморфозы, от адаптации к своеобразному у хордовых способу питания путем фильтрации, что, по классификации И. И. Шмальгаузена, должна быть отнесено к теломорфозам.
Точно так же у цеетод (другой "классический" пример ката- морфоза) катаморфные адаптации, обусловленные упрощением экологической обстановки - существование в кишечнике хозяина, не отделимы от теломорфозов - системы крючьев и присосок с управляющими этими прикрепительными органами мышцами. Для эктопаразитов, например вшей или пухоедов, подобное разграничение катаморфозов и теломорфозов вообще едва ли можно сделать. Возникает представление о катаморфозе как о крайнем выражении теломорфоза. Сужение адаптивной зоны - это всегда определенная степень упрощения отношений со средой и стабилизация этих отношений, что свойственно и теломорфозу (см. выше). Поэтому катаморфоз представляется не более чем крайним выражением этих свойств специализации как направления эволюции.
Вместе с тем представление о катаморфозе как о специализации, связанной с упрощением организации - общей дегенерацией, нуждается в уточнении. Это положение трудно применимо к паразитическим простейшим. Переход к паразитизму у жгутиковых и инфузорий часто связан не с упрощением, а с усложнением орга-низации, интерпретируемым как ароморфоз (Догель, 1954; Полянский, 1970, 1972). Подобная ситуация вполне объяснима именно для одноклеточных организмов, для которых химическая среда является основным комплексом факторов, требующих адаптации. Химизм же среды для паразитических форм несравненно сложнее, чем для свободно живущих, прежде всего благодаря большему разнообразию биологически активных веществ внутри организма-хозяина, чем в естественных водоемах или в почвенной воде. В этом своеобразии реакция адаптации простейших к условиям паразитизма состоит одно из выражений своеобразия организации простейших как клеток-организмов (Полянский, 1970, 1972).
Вторым аспектом эндопаразитизма, не укладывающимся в рамки представления о катаморфозе как об упрощении организации и свойственным не только Protosoa, но и Metasoa, является связанное со сменой хозяев усложнение жизненных циклов паразитических форм. Сложность перестройки организации по мере прохождения фаз жизненного цикла позволяет сравнивать сложность организации паразитических форм, например цестод (Гуляев, 1982), со сложностью свободно живущих форм.
Таким образом, упрощение организации при катаморфозе касается лишь морфологии стадий онтогенеза, но не преобразований ее при смене стадий. Даже для морфологии характерно сочетание собственно "катаморфных" признаков - признаков упрощения организации, и "теломорфных" признаков - приспособлений к существованию в условиях паразитизма или прикрепленного образа жизни. Причем следует учитывать, что эти морфогенетические преобразования не отражают еще мало исследованных биохимических изменений, без которых эндопаразиты не могли бы существовать в биохимически активной среде, каковой является внутренняя среда их хозяина. Едва ли к этим изменениям применимо представление о вторичном упрощении организации.
Еще более сложной оказывается организация паразитических форм, если рассматривать не только дефинитивную стадию их развития, а весь жизненный цикл. Сложнейшие приспособления промежуточных стадий онтогенеза, служащие для инвазирования промежуточных хозяев, для переживания периодов пребывания в среде между этапами инвазирования, преобразования организации, связанные с изменениями положения стадий в теле хозяина и т. д., нельзя рассматривать как катаморфозы - упрощения организации. Это теломорфные признаки. Следовательно, катаморфоз оказывается лишь частным случаем теломорфной специализации, или, что точнее, крайним выражением тенденции к упрощению организации, свойственной специализации, как направлению эволюции.
2.6.1.4. Гипоморфоз
Гипоморфоз - явление недоразвития, выражающееся в неотении, фетализации, педоморфозе (de Beer, 1930) - словом, в выпадении дефинитивных стадий онтогенеза всего организма или отдельных его частей, И. И. Шмальгаузен (1982) первоначально рассматривал как частный случай катаморфоза. Позднее он выделил гипоморфоз в качестве самостоятельного пути специализации (Шмальгаузен, 1969).
Для гипоморфоза, как и для любого другого типа специализации, характерно сужение адаптивной зоны. Однако оно связано в данном случае с этапностью онтогенеза. При эволюции по пути неотении происходит выпадение дефинитивной стадии онтогенеза и вместе с ним утрата всей той части экологической ниши вида, в которой обитала эта стадия. Наглядным примером сужения адаптивной зоны при гипоморфозе может служить неотения у хвостатых амфибий. С переходом к размножению на личиночной стадии развития и выпадением стадии взрослого организма Рrоteidae, Ambystomidae, Cryptobranchidae и др. формы утрачивают способность существовать на суше, сохраняя лишь "водную" часть своей адаптивной зоны. Таким образом, гипоморфоз с точки зрения экологии - это, несомненно, специализация. С катаморфозом, как подчеркивал И. И. Шмальгаузен (1982, 1969), его роднит упрощение организации: выпадение дефинитивной стадии онтогенеза влечет за собой генерализацию и упрощение морфологии, так как утрачиваются наиболее поздние филэмбриогенезы, создающие наиболее специальные адаптации взрослого организма (Северцов, 1981а). Поэтому, как было отмечено и де Бэром (de Beer, 1930), И. И. Шмальгаузеном (1982, 1969), гипоморфоз, будучи экологически специализацией, морфологически представляет собой деспециализацию - утрату наиболее специальных черт организации. На этом представлении основана теория педоморфоза (de Beer, 1930, Garstang, 1922), с которой полностью согласился И. И. Шмальгаузен (1982, 1969).
Согласно теории педоморфоза выпадение дефинитивных стадий онтогенеза создает предпосылки для резкого изменения направления эволюции, в результате чего могут возникать новые большие таксоны. Так, от олигомерных личинок многоножек путем педоморфоза, по-видимому, возникли насекомые (de Beer, 1930), от личинок асцидии таким же путем возникли аппендикулярии (Ливанов, 1955). Согласно Л. Больку (цит. по Шмальгаузену, 1969), явления фетализации имели место в становлении облика Homo spaiens. Строение черепа и других частей скелета человека грацильно по отношению к скелету человекообразных обезьян и напоминает морфологию их детенышей. М. А. Шишкин (1970) показал, что фетализация играла существенную роль в формировании отряда Anura. Таким образом, педоморфоз достаточно широко распространен в филогенезе, и через него могут возникать таксоны надвидового ранга, в том числе и ароморфные (см. ниже).
С теорией педоморфоза связано одно из остроумных объяснений неполноты палеонтологической летописи - наличие систематических разрывов между основаниями филогенетических рядов больших таксонов. Действительно, если выпадают дефинитивные стадии развития, то сравнительный ряд взрослых организмов, связывающий два таксона, построить невозможно просто в силу отсутствия этих организмов. Это не означает скачкообразности эволюции - она идет постепенно и непрерывно, но за счет формирования неотении и последующего возникновения новой дефинитивной стадии перерыв палеонтологической летописи становится принципиально невосполнимым. Второе объяснение таких перерывов связано, как сказано выше, с теорией квантовой эволюции.
Вместе с тем представление о педоморфозе противоречит на первый взгляд представлению об адаптивности промежуточных стадий онтогенеза. Действительно, утрата в ходе педогенеза дефинитивной стадии не означает утраты приспособлений личиночной стадии. Напротив, эти приспособления должны в ходе педогенеза прогрессивно развиваться, обеспечивая приспособление неотеничных форм к жизни в свойственных им условиях среды. Поэтому "генерализация" может коснуться лишь тех сторон организации гипоморфных форм, которые составляют адаптации взрослого организма и не функционируют на промежуточных стадиях онтогенеза. С точки зрения организма в целом утрата дефинитивных стадий развития таких частей едва ли будет представлять собой дискоординацию организации, так как провизорные адаптации и формируются в ходе эволюции в качестве приспособлений, обеспечивающих возможность дальнейшего онтогенеза дефинитивных признаков, т. е. координированность организации личинки формируется без этих признаков. Поэтому можно думать, что ни специализированность провизорных признаков, становящихся у неотеничных форм дефинитивными, ни координированность организации неотеников, по сути дела, не ниже, чем у форм, эволюция которых идет любым другим путем адаптациоморфоза. Иными словами, сама по себе неотения не ведет к педоморфозу. Она лишь обусловливает возможность этого способа эволюции. Возникновение же педоморфоза возможно, по-видимому, лишь при соблюдении условий, позволяющих занять новую адаптивную зону любому специализированному таксону, т. е. при ранней специализации в пограничной субзоне.
Действительно, ни среди рецентных, ни среди ископаемых неотеничных амфибий не возникли формы, которые эволюционировали бы по пути педоморфоза. Очевидно, этому препятствовала занятость соседних адаптивных зон, но одновременно это означает, что освоение новых зон наиболее вероятно в том случае, если они свободны, и что сам по себе педогенез является в лучшем случае предпосылкой педоморфоза, но не непосредственной его причиной (Симпсон, 1948).
Следует отметить, что гипоморфоз, как и другие формы специализации, редко выступает в чистом виде. Возникновение неотении связано, как правило, с дальнейшим приспособлением неотеничных форм к той среде обитания, в которой они существуют. Это приспособление может представлять собой не только специализацию, а также алломорфные и ароморфные адаптации. Достаточно ярким примером сочетания гипоморфоза с теломорфозом может служить организация европейского протея (Proteus anguineus). Эта неотеничная форма, полностью утратившая способность к метаморфозу, сохраняет в течение всей жизни признаки, свойственные личинкам хвостатых амфибий: наружные жабры, жаберные щели, личиночное строение гиобранхиального аппарата (Северцов, 1974), плавниковую складку на хвосте и ряд других признаков. Вместе с тем Proteus anguineus в отличие от второго рода и вида этого семейства Necturus maculatus приспособился к жизни в пещерах, поэтому вся его организация приобрела ярко выраженный "теломорфный" облик. Глаза протея рудиментарны, пигментация утрачена, третья жаберная дуга редуцирована (признак продвинутости по отношению к другим личинкам Urodela), сильно развиты мягкие губы, богато инервированные осязательными нервами. У Necturus maculatus, формы, живущей в водоемах на дневной поверхности, все эти признаки отсутствуют.
Организация аппендикулярий дает другой пример теломорфной эволюции гипоморфных форм. У этих животных сохраняются все признаки хордовых, свойственные личинкам асцидий. Однако фильтрационное питание в условиях планктонного образа жизни привело к сложной перестройке главной оси симметрии - хвост аппендикулярий подогнут концом вперед и повернут на 90° вокруг продольной оси, так что плавниковая складка лежит горизонтально. Кроме того, аппендикулярии строят вокруг своего тела домик, снабженный двумя отверстиями, затянутыми ловчими сетями.
Сочетание гипоморфоза и гиперморфоза среди современных форм дает Cryptobranchidae - гигантские саламандры, как и другие неотеничные Uredela, утратившие способность к обитанию вне воды. Среди стегоцефалов большинство неотеничных форм являются примерами сочетания гипоморфной и гиперморфной, а иногда- гипоморфной, гиперморфной и теломорфной специализаций. Примером таких форм могут служить Nectridia.
Алломорфные преобразования неотеничных форм, по-видимому, наиболее ярко выражены при возникновении травянистой растительности. Аналогичный пример дает фетализация, связанная с возникновением Anura, выразившаяся в редукции окостенений осевого скелета и частичной замене их хрящевыми образованиями и ряде других признаков (Шишкин, 1970).
Наконец, возникновение ароморфной организации на основе гипоморфоза может быть проиллюстрировано возникновением насекомых (в том случае, если их становление происходило так, как это представлял себе де Бэр) или становлением позвоночных (если оно происходило путем педоморфоза от личинок оболочников, как считал H. А. Ливанов, 1955). Наконец, фетализация предков человека служила основой эпиморфоза (Шмальгаузен, 1982).
Таким образом, гипоморфоз (фетализация) представляет собой основу для весьма разных направлений эволюции и в этом смысле организация гипоморфных форм не менее гетерогенна, чем других типов специализации. Однако, с другой стороны, гипоморфоз представляет собой, по-видимому, наиболее четко ограниченный тип специализации, не имеющий непрерывного спектра переходов, связывающего его с другими типами этого направления биологического прогресса. Своеобразие гипоморфоза обусловлено прежде всего тем, что он является результатом не столько дальнейшей адаптации к определенным экологическим условиям - сужение адаптивной зоны, сколько результатом утраты части зоны, свойственной предкам, или, что то же самое, гипоморфоз - не перестройка дефинитивной стадии развития, а ее утрата, т. е. специализация за счет промежуточных стадий онтогенеза предков.
Из приведенной характеристики форм специализации в рамках классификации И. И. Шмальгаузена (1982, 1969) следует, во-первых, что сами формы специализации гораздо более многообразны, чем те 4 типа, которые он выделил; во-вторых, все эти формы (за исключением гипоморфоза) плавно переходят друг в друга; в-третьих, они самым причудливым образом сочетаются друг с другом в течение онтогенеза данного организма и (или) в организации одной дефинитивной стадии онтогенеза специализированных форм. Иными словами, типы, выделенные И. И. Шмальгаузеном, сколь бы ни были привычны и удобны для характеристики некоторых форм специализации, недостаточны для характеристики всего разнообразия этого явления.
Видимо, для систематизации путей специализации можно было бы воспользоваться разработанной в экологии системой характеристик стенобионтности организмов по отношению к различным факторам среды: стенотермные, стеногалинные, стенотопные, стенофагические и т. д. формы. Однако такой подход не учитывает преобразований организации, связанных со специализацией, а именно анализом этих преобразований и ценна классификация
И. И. Шмальгаузена. Так, например, гипоморфоз и, видимо, гиперморфоз не укладываются в экологическую классификацию.
Строго говоря, классификация типов специализации должна учитывать три параметра: характер тех факторов среды, к которым приспосабливаются специализирующиеся формы, т. е. экологическую сторону специализации, характер преобразований организации, т. е. морфофизиологическую сторону специализации, и временную характеристику - степень специализации, точнее, степень продвинутости данной формы по пути данного направления специализации, по отношению к предкам. Подобная характеристика, по-видимому, достаточно полно может характеризовать любой случай сужения адаптивной зоны в процессе биологического прогресса специализирующихся форм. Однако такая трехмерная классификация представляется крайне громоздкой. Она, видимо, еще более сложна, чем система как таковая, а потому неприемлема из чисто практических соображений. На данном этапе развития наших знаний о главных направлениях эволюции создание систем классификации форм специализации едва ли оправдано. Там, где это возможно и удобно, следует, вероятно, пользоваться формами, выделенными И. И. Шмальгаузеном, сознавая при этом всю их относительность и неполноту.
Специализация в целом представляется именно единым направлением эволюции, связанным с сужением адаптивной зоны, постепенным замедлением темпов эволюции и стенобионтностью. Однако специализация далеко не обязательно влечет за собой вымирание. Биологический регресс и вымирание специализированных форм будут происходить только в том, хотя и очень распространенном случае, когда меняется сама адаптивная зона. Иными словами, маловероятно вытеснение специализированных групп конкурентами, находящимися на том же уровне организации. Палеонтологический материал позволяет показать смену низкоорганизованных форм более высокоорганизованными, но это не всеобщее явление. Часто специализация, при сохранении условий среды, к которым приспособлены специалисты, обеспечивает их неопределенно долгое существование. Особенно наглядно это видно на примере седентарных форм и паразитов. Весь арсенал современной медицины не способен пока еще обеспечить девостацию (полное уничтожение видов) и экто- и эндопаразитов человека. Уничтожение в конце 70-х гг., по-видимому, окончательное черной оспы явилось одной из величайших побед медицины. Наконец, именно специализированные формы дают начало новым ароморфозам.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'