Глава 18. Эволюционная палеонтология. (А. Н. Иванов)
К началу XX в. палеонтология достигла значительных успехов в познании истории органического мира на Земле. В главных чертах вырисовалось древо развития жизни, корни которого уходят в докембрийское время, а ствол и мощная крона простираются в палеозойской, мезозойской и кайнозойской эрах. Но в свете эволюционного учения были тщательно изучены лишь отдельные его ветви и побеги.
Перед палеонтологами XX в. стояла задача продолжить изучение филогении всех организмов, способных оставлять следы в слоях земли. Задача эта после Ч. Дарвина определилась как главная. По словам Л. Долло, она остается вечной задачей палеонтологии.
В палеонтологических данных по-прежнему нуждалась геология. Без них не может развиваться стратиграфия, с которой непосредственно связаны тектоника, палеогеография, геологическое картирование, а значит и планомерные поиски полезных ископаемых. Организуя палеонтологические исследования, геологические учреждения, конечно, подчиняли их в какой-то мере своим интересам и задачам. Но теоретические (в частности, филогенетические) исследования палеонтологов оказались необходимыми и здесь, способствуя усовершенствованию методов биостратиграфии.
В течение XX в. палеонтология не испытывала таких крупных преобразований, как в XIX в. Трудно ожидать их и в ближайшем будущем; это, однако, не означает, что палеонтология остановилась в своем развитии. Накапливались новые факты, вскрывались новые закономерности, возникали новые проблемы и направления исследований. Одни из них выдвигались логикой развития самой палеонтологии, другие возникали под влиянием прогресса в биологии и геологии.
Идеалистические концепции эволюции и дарвинизм
В начале XX в. кризис естествознания выразился в палеонтологии в возникновении виталистических антидарвинистских теорий эволюции. Они явились своеобразным преломлением успехов экспериментальной генетики и того факта, что палеонтология, против ожидания, не стала простым собранием иллюстраций к эволюционной теории Дарвина (см. также главу 17). Открывались новые факты и закономерности, которые, казалось, противоречили классическому дарвинизму.
Популярные у многих палеонтологов ламаркистские представления под влиянием успехов генетики терпят крах. Естественный отбор случайных, ненаправленных наследственных изменений также казался многим палеонтологам малоэффективным. Они считали эволюцию строго направленным, внутренне регулируемым процессом. Так ее представляли крупнейшие палеонтологи первой трети XX в.: О. Абель - в Европе, Г. Осборн - в США.
Венский палеонтолог Абель, известный своими важными палеобиологическими трудами, считал, что решающее значение в эволюции имеет сформулированный им "закон инерции органического развития" (1928). Сила "инерции" заставляет организмы изменяться строго прямолинейно в направлении специализации. Инерция действует и после того, как достигнуто приспособление к среде; тогда она приводит к бесполезному и даже вредному переразвитию специализированных признаков, и зашедшая в тупик филетическая ветвь вымирает. Абель превратил закон необратимости эволюции Долло в некий мистический принцип ортогенетической эволюции.

Генри Фейрфилд Осборн (1857 - 1953)
Осборн внес большой вклад в изучение эволюции млекопитающих, особенно носорогов и хоботных (1929 - 1942). Первоначально в своих теоретических взглядах он склонялся к ламаркизму. Но во второй половине жизни под влиянием идей А. Вейсмана о независимости зародышевой плазмы от сомы Осборн разочаровался в ламаркизме и выдвинул сложные, запутанные автогенетические теории, в которых витализм переплетался с энергетизмом. Его последним словом в области эволюционной теории было учение об "аристогенезе". "Аристогенез", по Осборну, есть "совершенно необъяснимый и таинственный" творческий процесс создания новых биомеханизмов. Направление его строго предопределено в наследственной субстанции и выдерживается вопреки влиянию среды. Развивающиеся признаки вначале совершенно бесполезны и становятся адаптивными лишь со временем.
В 20-х годах в Советской России появились две оригинальные теории эволюции, имевшие целью опровергнуть и заменить будто бы устаревший дарвинизм. Автором одной из них был известный зоолог и географ Л. С. Берг. В книге "Номогенез или эволюция на основе закономерностей" он, широко используя палеонтологический материал, стремился представить эволюцию как предопределенный процесс, протекающий под действием автономных, независимых от среды причин. Филогенез он считал строго закономерным и в этом отношении подобным онтогенезу.
Другая теория принадлежала геологу и палеонтологу Д. Н. Соболеву, взгляды которого во многом сходны со взглядами Берга. По мнению Соболева (1924), теорию Дарвина нельзя признать эволюционной теорией. Истинная эволюционная теория, утверждает Соболев, например ламаркизм, предполагает "развитие начал, заложенных в самом развивающемся существе", а у Дарвина есть лишь "механическое сложение независимых органических изменений"*, Соболев считал, что эволюция "не зависит от игры случая", которой пользуется отбор, а происходит направленно по независимым от отбора законам. В противоположность дарвинистам, Соболев и Берг считали эволюцию процессом полифилетическим.
* (Д. Соболев. Начала исторической биогенетики. Гос. изд-во Украины, 1924, стр. 32.)
Взгляды Берга и Соболева встретили решительные возражения советских биологов и не оказали заметного влияния на отечественную палеонтологию. Более того, советские палеонтологи взяли на себя задачу защиты и дальнейшего развития дарвинизма. Среди них особое место принадлежит А. А. Борисяку, который относил к числу важнейших задач эволюционной палеонтологии выяснение путей и закономерностей эволюции с помощью исследования конкретных филогенезов, изучение взаимоотношения организмов и среды и проблему видообразования.
На долю советской науки выпала задача критической оценки с позиций диалектического материализма того теоретического багажа, который накопился в палеонтологии после Дарвина. Эта важная для современной палеонтологии работа получила наиболее полное выражение в книгах Л. Ш. Давиташвили "Развитие идей и методов в палеонтологии после Дарвина" (1941) и "История эволюционной палеонтологии от Дарвина до наших дней" (1948).
Определение возраста Земли
В начале XIX в. Ж. Кювье в соответствии со священным писанием укладывал историю Земли в 7 тыс. лет. В конце того же века по подсчетам физика В. Томсона возраст Земли как остывающего тела определялся в 100 млн лет.
После открытия в 1896 г. французским физиком А. Беккерелем радиоактивного распада и последующего изучения радиоактивных веществ возникла возможность выяснить абсолютный возраст горных пород, содержащих продукты распада. На эту многообещающую возможность указал уже английский геолог Д. Джоли в книге "Радиоактивность и геология", вышедшей в 1909 г. А в 30-х годах появились расчеты длительности геологических эр и периодов в миллионах лет. Палеонтологи получили возможность судить о скорости эволюции в разные периоды истории Земли. Одной из древнейших пород была признана порода из Карелии с абсолютным возрастом в 1600 млн лет, что позволило определить общий возраст Земли в 3 млрд лет. Но в середине 60-х годов были открыты горные породы с абсолютным возрастом 3,5 млрд лет. Общий возраст Земли определяется теперь по крайней мере в 4,5 млрд лет, а следы жизни известны из архейских слоев с возрастом около 3 млрд лет.

Алексей Алексеевич Борисяк (1872 - 1944)
Успехи в изучении палеонтологической летописи
В 1940 г. И. А. Ефремов объединил все направления по изучению процессов захоронения органических остатков в единую отрасль и предложил именовать ее тафономией. Установив условия захоронения и сохранения древних форм животных и растений и определив, в каких местах их обнаружение маловероятно (как, например, в горных районах), тафономия в немалой степени способствовала успешной деятельности палеонтологических экспедиций.
Познание палеонтологической летописи связано прежде всего с открытием и описанием новых видов ископаемых организмов. Но рост общего числа известных ископаемых форм шел также за счет разделения и повторного описания ранее описанных видов. Для XX в. характерны стремление к упорядочению таксономии, к унификации описании и привлечению для определения ископаемых ЭВМ, а также возрастание интереса к вопросам теоретической систематики. Пятитомная сводка по ископаемым организмам всего мира, законченная немецким палеонтологом К. Циттелем в 90-х годах прошлого столетия, за полвека настолько устарела, что в 50-х годах нашего века началось издание новых сводок: по палеозоологии - во Франции под редакций Ж. Пивто, по беспозвоночным - в США под редакцией Р. Мура и по ископаемым организмам в целом - в СССР под редакцией Ю. А. Орлова (Ленинская премия, 1967). Монументальный коллективный труд советских палеонтологов - "Основы палеонтологии" в 15 томах (1954 - 1964) - полный справочник о родах ископаемых животных, растений и протистов.
К числу достижений в прочтении палеонтологической летописи относится также уточнение представлений о давно известных скелетных образованиях и их биологическая расшифровка. Это стало возможным благодаря новым находкам, совершенствованию методов исследования и теоретическому осмысливанию ранее известных фактов.
Так, например, известные палеозойские колониальные морские организмы с роговым скелетом - граптолиты обычно причислялись к типу кишечнополостных, В 1938 г. польский палеонтолог Р. Козловский доказал близость граптолитов к птеробранхиям, о чем предположительно высказывался еще в 1905 г. А. Щепотьев. В результате граптолиты в качестве особого подтипа были отнесены к типу полухордовых. Уникальная коллекция, собранная в Поволжье, в районе Ульяновска, позволила К. А. и Г. К. Кабановым (1959, 1967) с уверенностью заявить, что ростр белемнитов был первоначально мягким образованием и обызвествлялся только вторично. Л. Ш. Давиташвили (1961) показал, что загадочные структуры у динозавров и других ископаемых форм, известные под названием "эксцессивных образований", получают объяснение на основании теории полового отбора как перигамические признаки.
Новые методы стратиграфии
Интенсивное развитие нефтяной промышленности и большой размах геологоразведочных работ по нефти и горючим газам, сопряженный с глубоким бурением, дающим в руки стратиграфа обычно только микрофауну, вынудило палеонтологов обратиться к изучению фораминифер. Постепенно фораминиферы приобрели решающее значение в стратиграфии толщ морского происхождения и стали основным объектом микропалеонтологии. К этому разделу палеонтологии отнесли также исследование радиолярий, спор и пыльцы растений, а также диатомовых водорослей. Анализ спор и пыльцы приобрел значение основного метода в стратиграфии толщ континентального происхождения и вошел в науку под названием споро-пыльцевого анализа. Выяснение процентного отношения между количеством спор и пыльцы разных видов в серии последовательных образцов дает материал для пыльцевой диаграммы, которая позволяет делать заключение об изменении климатических условий и растительности в данном районе.
Основателем метода споро-пыльцевого анализа считается шведский торфовед Л. Пост. В начале 90-х годов XIX в. он стал выражать соотношение пыльцы разных видов в процентах, а в 1916 г. применил пыльцевые диаграммы. В СССР развитие метода споро-пыльцевого анализа связано с именем В. Н. Сукачева, опубликовавшего первую работу о составе пыльцы в торфе еще в 1906 г. Первоначально этот метод применялся к исследованию торфов современной эпохи, затем погребенных торфяников плейстоцена и, наконец, древних континентальных толщ.
Широкое применение споро-пыльцевого метода отразило тенденцию палеонтологии к переходу от изучения истории отдельных групп к изучению истории флор и фаун. Отвечая практическим запросам статиграфии и палеонтологии, эта тенденция вместе с тем отражает и сдвиги, происходящие в биологии,- стремление к изучению биогеоценозов. Кроме того, как палеонтологи, так и геологи разными путями приходят к необходимости всестороннего исследования скелетных остатков и следов жизнедеятельности ископаемых организмов вместе с заключающим их слоем. Геологи ставят во главу угла восстановление среды образования осадков и называют такое исследование биофациальным анализом. В палеонтологии для этого применяются разные термины. Абель предпочитал термин палеобиологии. В СССР в 30-х годах Р. Ф. Геккером введен в обращение термин палеоэкология.
В 20-х годах В. И. Вернадский разработал учение о биосфере, огромное значение которого в наше время признается как для наук о Земле, так и естествознания в целом (см. также главы 17 и 27). Биосфера, но Вернадскому,- это земная оболочка, населенная организмами и характеризующаяся сложным взаимодействием живого и неживого. Слои осадочных горных пород с превосходно сохранившимися остатками организмов и лежащие глубже метаморфизированные породы докембрия с трудно различаемыми следами жизни являются продуктами биосферы, созданными в разное время. В настоящее время палеонтологи и геологи имеют возможность вести комплексные исследования населения водоемов и материков в его взаимодействии со средой, т. е. изучать историю биосферы в целом.
Новые находки - вклад в филогению
Достижения палеонтологии XX в. в исследовании филогении органического мира в немалой мере зависели от новых находок и открытий. Благодаря изучению нижнедевонской и верхнесилурийской флоры псилофитов, начатому в 20-х годах, был решен важнейший вопрос истории растительного мира. Псилофиты - первые сосудистые растения суши. От них произошли папоротники, хвощи, плауны и мхи. Первая находка псилофитов была описана в Америке еще в 1859 г. Но только в XX в. с новыми находками в Европе и их правильной оценкой был заполнен зияющий пробел в геологической истории растений. Псилофиты оказались промежуточной группой между водорослями и высшими наземными растениями.
Не меньшее значение имело открытие и описание шведскими палеонтологами Э. Стеншьо, Г. Севе-Седербергом, Е. Ярвиком в начале 30-х годов в верхнедевонских слоях Гренландии скелетов древнейших наземных позвоночных животных. В капитальном труде И. И. Шмальгаузена (1964), посвященном проблеме происхождения наземных позвоночных животных, примитивные амфибии из верхнего девона Гренландии (ихтиостеги) охарактеризованы как недостающее звено в эволюции от рыб к наземным четвероногим, поскольку по строению, образу и времени жизни они оказались настоящими переходными формами между кистеперыми рыбами и амфибиями. Выход позвоночных на сушу и дальнейшая эволюция фаун наземных позвоночных в истории Земли связаны с эволюцией насекомых, которые служили им важнейшим источником белковой пищи. На это обратил внимание Б. Б. Родендорф (1970).
Из новых местонахождений костей наземных позвоночных особую известность приобрела Центральная Монголия. Здесь работали палеонтологические экспедиции США (1921 - 1930), СССР (1946 - 1949) и Польши (1962 - 1964). Они добыли большое число скелетов меловых ящеров и меловых и третичных млекопитающих, что способствовало изучению их эволюции, экологии и систематики.
Вопреки распространенному представлению о вымирании четырехлучевых кораллов в конце пермского периода из триасовых отложений Закавказья были описаны несколько семейств этой группы, в том числе плерофиллиды (Т. Г. Ильина, 1963), которые по целому комплексу признаков оказались переходными формами между четырехлучевыми и шёстилучевыми кораллами.
Выяснилось, что некоторые группы современной морской фауны, считавшиеся давно вымершими, существуют и ныне. В 1938 г. в Индийском океане, у юго-восточных берегов Африки, была впервые поймана кистеперая рыба, описанная под названием латимерии. В 1952 г. датской глубоководной экспедицией в Тихом океане, близ Мексики, был обнаружен моллюск из класса моноплакофор, названный неопилиной. Если кистеперые и моноплакофоры оказались "живыми ископаемыми", то морские донные глубоководные беспозвоночные животные - погонофоры, несмотря на свой "древний" облик, до последнего времени в ископаемом состоянии не были известны. А. В. Иванов (Ленинская премия, 1961), установивший в 1955 г. тип погонофор, не сомневался в возможности обнаружения ископаемых представителей этого типа (см. также главу 1). Это предвидение блестяще подтвердилось: Б. С. Соколов* обнаружил трубки погонофор в кембрийских слоях, выделив их в отряд сабеллитид (1965).
* (Б. С. Соколов. Древнейшие отложения раннего кембрия и сабеллитида.- Всемирный симпозиум по палеонтологии докембрия и раннего кембрия. (Тезисы докладов). Новосибирск, 1965.)
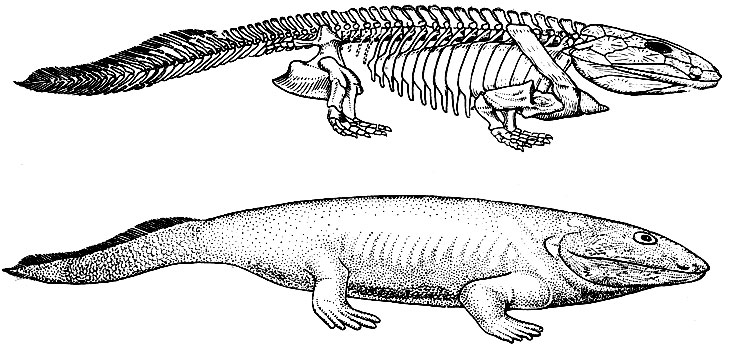
Древнейшие земноводные - Ichthyostega. Реставрация (по Ярвику, 1952)
Упомянутые находки ранее неизвестных ископаемых форм свидетельствуют о вероятности подобного рода открытий и в будущем.
Отсутствие переходных форм и сальтационные теории эволюции
Палеонтологическая летопись сохранила формы, дающие возможность проследить морфологические преобразования в пределах низших систематических подразделений (разновидность, подвид, вид, род, семейство). Известно немало эволюционных рядов таких форм. Попытки проследить документально возникновение более высоких систематических групп (отрядов, классов, типов) часто оказываются безуспешными ввиду того, что редкость нахождения промежуточных форм возрастает пропорционально повышению систематического ранга. Поэтому связи между крупными систематическими группами в филогенетическом древе часто остаются гипотетичными. В отсутствие палеонтологических находок они устанавливаются на основе данных сравнительной анатомии, эмбриологии и геологической истории. Так, например, неизвестны исходные формы покрытосеменных растений, и последние гипотетически выводятся из голосеменных. Остаются неизвестными переходные формы между пресмыкающимися и млекопитающими.

Скелет гигантского хищного динозавра Tarbosaurus efremovi Maleev, добытого в Монголии палеонтологической экспедицией АН СССР
В сводке по филогении беспозвоночных Д. М. Федотов* привел таблицу с изображением стволов, соответствующих восемнадцати типам. Они идут, не соединяясь между собой, от кембрия до современной эпохи. Только в докембрии гипотетически конструируется их ответвление от главного ствола. По мере изучения беспозвоночных число самостоятельных типов росло, нередко за счет классов, ранее относившихся к одному типу.
* (Д. М. Федотов. Эволюция и филогения беспозвоночных животных. М., "Наука", 1966.)
В палеонтологической летописи, как правило, не обнаружено промежуточных форм даже между классами одного и того же типа.
Тип, класс, отряд и надстройки к ним выражают существенные различия в плане строения организмов, тогда как низшие систематические категории соответствуют различиям, которые не затрагивают плана строения. Подмеченное еще до Дарвина отсутствие среди ископаемых организмов переходных форм между крупными систематическими подразделениями стало в последарвиновский период важнейшей проблемой эволюционной палеонтологии.
Отсутствие переходных форм между типами палеонтологи-антидарвинисты использовали в качестве главного козыря в борьбе с эволюционизмом. Так, немецкий палеонтолог Е. Дакке (1935) допускал эволюцию только в пределах типа организации. Другие палеонтологи разделяют эволюцию органического мира на принципиально различные, управляемые разными факторами фазы и проповедуют скачкообразное превращение одних типов в другие без переходных форм. Что касается эволюции, действующей в рамках типа, то она преобразует разновидности, виды и роды через переходные формы.
Немецкий палеонтолог О. Шиндевольф (1936 и позднее) называл решающую фазу эволюции, связанную с превращением типов организации, неогенезом или типогенезом, а свою концепцию эволюции - типострофизмом. По его мнению, крупные мутации резко преобразуют тип на ранней стадии онтогенеза и потому переходных форм вообще быть не может. Мутирование происходит лишь в эпохи повышенной радиации. Новый тип организации, возникнув скачком, преадаптивен, и организмы сами разыскивают подходящую себе среду. После взрывной фазы возникновения новых морфологических типов наступает фаза типостаза. Для нее характерно ортогенетическое приспособление на основе мелких мутаций под действием естественного отбора. При ней изменения происходят на поздней стадии онтогенеза и тип организации не затрагивается. Возникают эволюционные ряды форм, связанные между собой переходами. Позже Шиндевольф стал выделять еще фазу типолиза, или упадка в истории типа (чрезмерная специализация, вымирание).
Другой немецкий палеонтолог, К. Бойрлен (1937), подобно Шинде-вольфу, выделил две основные фазы эволюции. Превращение одного типа организации в другой, по Бойрлену, не может происходить путем мелких изменений на поздних стадиях онтогенеза, поскольку прежде должно быть утрачено специализированное, потерявшее эволюционную пластичность состояние. Это достигается путем "неоморфоза", под которым Бойрлен понимает остановку развития на той стадии онтогенеза, которая предшествует формированию признаков старого типа. Эти признаки как бы отбрасываются, происходит деспециализация и открываются возможности для формирования признаков нового типа. В связи с этим Бойрлен считал, что нет надобности искать каких-то неспециализированных предков и переходные формы между разными типами организации. Организмы всегда были в той или иной мере специализированными, но благодаря неоморфозу специализация не мешает эволюции. Последняя - "ортогенетическая" - фаза эволюции осуществляется на основе приспособительных изменений на конечной стадии онтогенеза. Она удлиняет онтогенетический цикл, ведет к специализации и старению филетической ветви. Только неоморфоз может открыть новый путь эволюции.
В отличие от автогенетических построений Шиндевольфа и Бойрлена американский палеонтолог Дж. Симпсон (1944) сделал попытку истолковать отсутствие переходных форм между высшими систематическими категориями с позиции синтетической теории.
Симпсон различал три формы эволюции: видообразование, доступное изучению генетиков; филетическую эволюцию, происходящую на уровне рода и семейства и прослеживающуюся палеонтологами в виде линейных рядов типа ряда лошадей; квантовую эволюцию, которая за короткое время приводит к коренным изменениям структурных и физиологических систем и появлению новых семейств, отрядов и классов. Особо крупные сдвиги, порождающие новые классы и типы, он обозначал термином мегаэволюция. При квантовой эволюции и тем более мегаэволюции переходные формы отсутствуют, но не потому, что их не было. Они существовали недолго и на ограниченной территории, а потому их остатки редко сохранялись.
Все формы эволюции, по Симпсону, основываются на мутациях. Представление о квантовой эволюции он связывал с допущением крупных мутаций или повышенного темпа мутирования под действием повышенной радиации. Но эти предположения пока остаются не доказанными. Симпсон признавал также роль генетического дрейфа (генетико-автоматических процессов). Наиболее благоприятными условиями для возникновения признаков новой организации, по Симпсону, служит малочисленность и полная изоляция популяций.
Соотношение филогенеза и онтогенеза в свете палеонтологических данных
Возникновение переходных форм в эволюции прежде всего зависит от способов преобразования онтогенеза. Поэтому проблема переходных форм связана с проблемой исторической взаимозависимости между онтогенезом и филогенезом.
Критический анализ воззрений Э. Геккеля на взаимоотношения онтогенеза и филогенеза основывалась как на детальных сравнительно-эмбриологических работах, так и на палеонтологических данных.
При исследовании онтогенеза раковины в некоторых эволюционных рядах ископаемых головоногих моллюсков - аммонитов обнаружились отношения, обратные тем, которые следовало ожидать, исходя из биогенетического закона. Признаки средних оборотов раковины оказались сходными с таковыми у взрослых потомков. А. П. Павлов (1910) назвал упомянутое явление "пророческими", или "профетическими", фазами, Д. Н. Соколов - "вставными фазами" (1912), Л. С. Берг - "филогенетическим ускорением" (1922), а О. Шиндевольф - "протерогенезом" (1925). В представлениях Шиндевольфа протерогенез занимает видное место как особый модус раннеонтогенетического становления нового типа. При эволюции по этому модусу вновь возникший тип выражен только в онтогенезе, во взрослом же состоянии форма возвращается к типу предка. Только постепенно в ходе филогенеза новый тип выявляется и во взрослом состоянии. Шиндевольф иллюстрирует это эволюционными рядами аммонитов, фораминифер, кораллов и даже человека.
Наблюдая необъяснимые при помощи биогенетического закона явления и не удовлетворяясь их толкованием в духе "протерогенеза", многие западноевропейские палеонтологи (например, знаток мезозойских аммонитов Л. Спат) стали игнорировать изучение онтогенеза при установлении филогенетических отношений у аммонитов. В советской палеонтологии исследование онтогенеза в целях выяснения филогенеза и построения естественной системы продолжалось. В. Е. Руженцову (1960) удалось показать, например, на палеозойских аммонитах, что эволюция их раковины, в частности лопастной линии, шла, как правило, по способу анаболии, допускающей рекапитуляцию.
При изучении соотношения онтогенеза и филогенеза в классических рядах с так называемыми "профетическими фазами" Павлову удалось показать, что явления, обозначенные этим термином, получают материалистическое, дарвинистское истолкование (А. Н. Иванов, 1945). Преходящие в онтогенезе стадии могут у потомков распространяться на взрослое состояние (брадигения), а в других случаях, наоборот, признаки взрослого состояния - на более раннюю стадию (тахигения). И то, и другое может происходить под действием естественного отбора. В популяции, кроме форм, имеющих нормальную скорость онтогенеза, обнаруживаются особи, раньше и позже вступающие в конечную стадию развития (тахиморфные и брадиморфные). В случае полезности замедленного развития отбор соответствующих мутаций обеспечит сохранение признаков преходящей стадии до взрослого состояния.
Значение неотении
Еще во второй половине XIX в. зоологи и палеонтологи указывали на эволюционное значение различного рода задержек и остановок в развитии, из-за которых признаки взрослого организма или органа не развиваются, и эта стадия как бы отбрасывается. Это явление называли по-разному. Особенно популярным оказался термин "неотения", предложенный Э. М. Кольманом в 1884 г. Классическим примером неотении является современный американский аксолотль - способная к размножению личинка амблистомы. Нечто подобное аксолотлю в палеонтологии представляет пермский лабиринтодонт двинозавр, открытый и описанный В. П. Амалицким в 1921 г.
Английские биологи В. Гарстанг в 1922 г. и Г. де Бер в 1930 г. под названием "педоморфоза" описали особый случай филогенетических изменений, выражающийся в остановке онтогенеза и последующих новообразованиях. По их мнению, этим способом в истории животного мира могли осуществляться превращения типов, классов, отрядов. Так, насекомые, по их мнению, произошли от личинок многоножек, хордовые - от личинок иглокожих. При этом специализация взрослых предков не мешала эволюции, так как взрослая стадия не достигалась. Значит, нет надобности всегда искать неспециализированного предка.
А. Н. Северцов и И. И. Шмальгаузен рассматривали задержки развития как формы регрессивной эволюции. Шмальгаузен (1939) считал, что постоянножаберные амфибии - это утратившие способность превращаться в наземное животное половозрелые личинки, что коловратки, пантоподы, клещи, мшанки могли возникнуть путем недоразвития. Недоразвитие, по его мнению, "быть может... действительно лежит в основе наиболее крупных преобразований организации"*.
* (И. И. Шмальгаузен. Пути и закономерности эволюционного процесса. М.- Л., Изд-во АН СССР, 1939, стр. 161.)
А. Л. Тахтаджян в ряде работ начиная с 1943 г. выяснил большое значение неотенических явлений в эволюции растительного мира, в частности, покрытосеменных. "Неотеническое преобразование коснулось, по-видимому, всех органов и частей предков покрытосеменных, что не могло не привести к определенному перерыву в ряду взрослых форм. Поэтому происхождение покрытосеменных было не только быстрым (результат эволюции на основе изолированных периферических популяций), но и прерывистым (результат неотении)"*. Такой важный шаг в эволюции жизни, как появление травянистых покрытосеменных, по мнению Тахтаджяна, также осуществился путем неотении. Травы возникли из деревьев путем "ярусной неотении".
* (А. Л. Тахтаджян. Происхождение покрытосеменных растений. М., Изд-во АН СССР, 1961, стр. 19. )
О простоте генетических механизмов неотении писали Дж. Холдейн (1932) и Н. К. Кольцов (1936). Они допускали, что мутации генов, контролирующих скорость онтогенеза, или даже мутация одного гена могут повлиять на все развитие. При неотении сохраняется богатый генотип предков, что благоприятствует последующей прогрессивной эволюции.
Кроме упоминавшегося выше динозавра палеонтология располагает многими другими примерами эволюции путем задержки развития, и их число непрерывно умножается. Недавно Д. В. Обручев (1969) обратил внимание на большое значение педоморфоза в эволюции низших позвоночных. М. А. Шишкин (1969) показал, что бесхвостые амфибии произошли от рахитомных лабиринтодонтов путем замедления развития, обусловившего сохранение в строении головы потомков личиночных черт предков. Вместе с тем М. А. Шишкин (1967) вновь поднял вопрос о возможности частичной обратимости эволюции. Появилось немало работ, демонстрирующих роль педоморфоза в эволюции беспозвоночных, в частности моллюсков и иглокожих. Все более выявляющееся значение педоморфоза в эволюции проливает свет на причины отсутствия переходных форм между крупными систематическими группами.
Проблема полифилетического происхождения таксонов
Известно, что Дарвин допускал только монофилетический путь эволюции, при котором новая группа появляется от одного корня (вида) и в одном географическом месте (районе). Противники Дарвина, а ныне и некоторые его сторонники допускают полифилию, т. е. развитие новой группы из многих корней в виде параллельных линий. При этом обсуждается возможность полифилетического возникновения всех ступеней жизни, от первичных организмов до человека. Вернадский в посмертно изданной работе писал: "Сейчас, мне кажется, можно твердо стоять на той точке зрения, что представление о едином предке - монофилетическое представление, или о немногих предках противоречит тому, что мы знаем о геологическом значении живого вещества"*. Не без влияния Вернадского изменилось представление о происхождении жизни: возникновение первичных организмов разных видов во многих географических районах пользуется теперь широким признанием.
* (В. И. Вернадский. Химическое строение биосферы Земли и ее окружения. М., "Наука", 1965, стр. 291.)
Непрерывно увеличиваются данные о полифилетическом происхождении таксонов. Это приводит систематиков, стремящихся к построению филогенетически обоснованной системы, к необходимости дробить привычные, широкие по охвату таксоны. Так, выяснилось, например, что класс млекопитающих происходит из многих корней. Классы представляют собой "уровни" организации и объединяют гетерогенные формы, вышедшие пучком из предшествующего уровня (класса). Симпсон, однако, не считает нужным дробление старого класса млекопитающих и предлагает считать "монофилетическим" всякий таксон, если доказано его происхождение от таксона того же ранга по одной или многим линиям. Так, все млекопитающие, поскольку они произошли от пресмыкающихся, остаются единым классом. Соображения Симпсона находят поддержку у Э. Майра и других ученых.
В последнее время усилился интерес палеонтологов к явлениям параллелизма, состоящим в том, что у близких по происхождению таксонов, например родов в пределах семейства, параллельно возникают сходные особенности, которых не было у их предка. Этот параллелизм в развитии гомологичных органов связан с параллелизмом мутаций, возникающих по закону гомологических рядов Н. И. Вавилова на основе близости генотипов, унаследованных от общих предков. Исследование взаимоотношения между явлениями полифилии и параллелизма в эволюции - одна из актуальных задач палеонтологии и эволюционного учения.
Изучение докембрийских организмов
Большое значение в палеонтологии XX в. приобрела проблема древнейших организмов. В конце прошлого века палеонтологи разных стран описали из докембрийских слоев остатки губок, кишечнополостных, червей, брахиопод, моллюсков, иглокожих и других беспозвоночных. При пересмотре этих находок в наше время обнаружилось, что одни из них - и таких большинство - представляют собой неорганические образования, другие отнесены к докембрию ошибочно. Споры наземных растений, открытые С. Н. Наумовой в конце 40-х годов, оказались вмытыми из вышележащих слоев. Теперь из собственно докембрия достоверно известны только остатки водорослей (в частности, широко распространенные известковые образования - стромотолиты и онколиты, созданные синезелеными водорослями), остатки бактерий, а также ряд микроскопических образований, напоминающих яйца и цисты мельчайших животных и споры морских растений. Большой вклад в изучение многообразных остатков водорослей из докембрийских слоев в СССР внес А. Г. Вологдин (1962).
Таким образом, проблема древнейших организмов стала вместе с тем проблемой перехода от низшей ступени развития жизни водоемов в докембрии (преимущественно предъядерные организмы) к сложному и богатому населению моря кембрия, включавшему все типы беспозвоночных. Прежние ссылки на метаморфизм докембрийских слоев утратили свою силу. Совершенствование методов исследования горных пород и химической препаровки позволяют обнаруживать ископаемые остатки микроорганизмов и выделять органические образования. Датировка слоев при помощи метода радиоактивного распада дает возможность подойти к выяснению этапов в истории докембрийской жизни.
Крупным событием в палеонтологии докембрия было открытие Р. Сприггом в 1947 г. и последующее детальное изучение М. Глесснером (1958 - 1965) ископаемых из песчаников Эдиакары в Южной Австралии. Описано свыше двух десятков видов, принадлежащих кишечнополостным, червям и каким-то предшественникам моллюсков и иглокожих. Все они лишены скелета и сохранились в виде отпечатков. Фауна Эдиакары и близкие ей фауны Южной Африки, Англии и СССР относятся к периоду между 670 и 550 млн. лет, т. е. предшествовавшему кембрию.
Отсутствие минерального скелета у докембрийских организмов и, напротив, его наличие у многих животных кембрия приводят к предположениям о существовании иных физико-химических условий в докембрии и иной физиологии докембрийских организмов.
Взаимодействие истории Земли и жизни
Уже второе столетие палеонтология и геология собирают материалы о взаимодействии между развитием органического мира и историей Земли. И все же имеющиеся знания очень далеки от полноты и ясности.
По современным взглядам, познание истории Земли и жизни в их связи и взаимной обусловленности в конечном счете позволяет реконструировать историю биосферы. Изменения земной коры (тектогенез) в своей основе не зависят от эволюции органической жизни, хотя жизнь оказала сильное влияние на геохимические процессы и осадочное породообразование. Как показал Н. М. Страхов (1936), в истории Земли ярко выступает прогрессирующее вытеснение хемогенной седиментации частично терригенной, частично биогенной седиментацией, т. е. первичные осадки преимущественно химической природы уступают место вторичным осадкам обломочного и биогенного происхождения. Влияние эволюции земной коры на эволюцию жизни гораздо глубже и многостороннее. Геологические изменения - первопричина того непостоянства физико-географических условий на поверхности Земли, без которого эволюция жизни невозможна. Было установлено, что крупные смены в составе растительного и животного населения суши совпадают с эпохами мощных горообразований, в эпохи же геологического "затишья" эволюция органического мира протекала относительно спокойно.
В самом начале XX в. Г. Осборн привлек внимание исследователей истории органического мира к закону "адаптивной радиации". Согласно этому закону, после возникновения нового типа организации (например, класса животных) происходит его расселение и приспособление к самым разнообразным условиям жизни. При этом возникают специализированные группы, генеалогически расходящиеся в разные стороны от общего ствола. Как показал Осборн на млекопитающих, в третичном периоде произошла радиация плацентарных и независимо от нее радиация сумчатых в Австралии, издавна изолированной от других материков.
Но адаптивная радиация была характерна не только для млекопитающих. На суше, как бы волнообразно, происходила смена господствующих групп животного и растительного царств. Так, в истории позвоночных четко выделяются следующие последовательно сменявшие друг друга "волны": стегоцефалы (карбон), звероподобные ящеры (пермь), мезозойские ящеры, млекопитающие (третичный период) и человек (антропоген).
Согласно теории естественного отбора, новая, более совершенная группа должна вытеснять старую. Смена же "волн" жизни в истории Земли не всегда укладывается в эту схему. Не всегда ясны отношения каждой последующей "волны" к предыдущей и к изменению физико-географических условий. Еще недостаточно изучены конкретные причины и обстоятельства, приводившие к вымиранию одних групп и к процветанию или вспышкам в развитии других.
В истолковании некоторых событий истории органического мира палеонтологи все чаще обращаются к влиянию космических агентов. О. Шиндевольф (1953 и позднее) отстаивает идею о большой роли периодического усиления радиации, поступавшей на Землю из космоса, которая, по его мнению, могла вызвать резкое увеличение частоты мутаций и ускорение хода эволюции. Другие исследователи непосредственно связывают с увеличением радиации такие события, как вымирание мезозойских динозавров. Не исключено, что в будущем будут найдены методы определения интенсивности радиации в разные геологические эпохи и будет выяснено ее конкретное влияние на ход эволюции органического мира. Так, по соотношению изотопов кислорода в раковинах морских беспозвоночных стало возможным судить о температуре воды в морях далеких геологических эпох.
Начальные этапы в развитии органического мира, несомненно, зависели от формирования в атмосфере озонового экрана, защищающего живое от губительного действия ультрафиолетовых лучей. Было установлено, например, что выход растений и животных на сушу мог произойти лишь после того, как в результате фотосинтеза водных автотрофов в атмосфере накопилось достаточное количество кислорода, а вместе с тем и озона. Как показал недавно Давиташвили*, представления о так называемых "фаунистических разрывах" во многом зависят от неточностей в геологической синхронизации исчезновения и появления сменявших друг друга групп организмов. Объективная оценка современных данных по этому вопросу убеждает в том, что основным фактором вымирания таксонов было воздействие со стороны новых конкурентов и новых хищников. Это справедливо и в отношении вымирания крупных пресмыкающихся на границе мезозоя и кайнозоя. Млекопитающие, по новым данным, были широко распространены уже в раннемеловое время. Вопросы вымирания древних рептилий исследовал также Л. К. Габуния (1969).
* (Л. Ш. Давиташвили. Причины вымирания организмов. М., "Наука", 1969.)
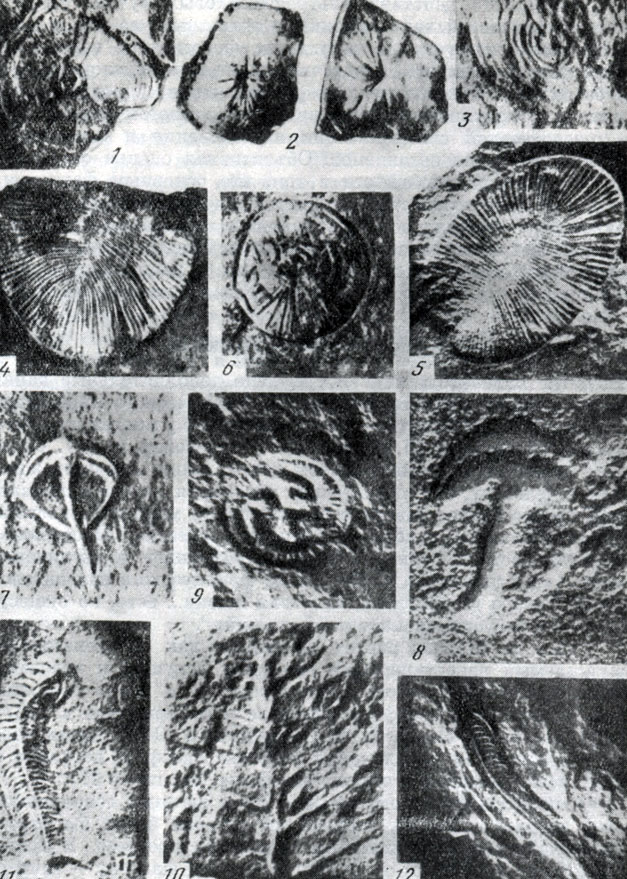
Отпечатки животных позднего докембрия в кварците. Эдиакары (Южная Австралия) (по Глесснеру, 1963): 1 - Cyclomedusa radiata Sprigg, медуза, X 3/8; 2 - Pseudorhizostomites sp., x 3/8; 3 - Spriggia annulata Sprigg, медуза, X 3/8; 4 - Dickinsonia costata Sprigg, червь, X 1; 5 - та же форма, X 1/2; 6 - Cyclomedusa inflata Sprigg, медуза, X 1/2; 7 - Parvancorina minchami Gl., червь X 3/4; 8 - та же форма, X 7/12; 9 - Tribrachidium heroldicum Gl., трибрахидиум, совершенно новая форма, не имеет аналогов среди ныне живущих организмов, х 3/4; 10 - Rangea arborea Gl., морское перо, X 3/4; 11 - Spriggina floundersi Gl., трилобитоподобный червь х 9/10; 12 - та же форма, х 15/16
Изучение прогрессивной эволюции
Развитие палеонтологии в XX в. дало дополнительные данные в пользу прогрессивного характера эволюции органического мира. Однако эти данные получили идеалистическое истолкование у многих палеонтологов-финалистов (Г. Осборн, Ш. Депере, О. Шиндевольф и др.). Сконцентрировав свое внимание на магистральном направлении эволюции животных, ведущем к человеку, они, сознательно игнорируя другие направления (например, прогрессивное развитие растительного мира и беспозвоночных), объявили эволюцию целенаправленным процессом. Прослеживая развитие нервной системы, они отмечают "стремление" к увеличению головного мозга ("цефализация"*). По словам французского палеонтолога и философа П. Тейара де Шардэна, "история жизни... есть, по существу, развитие сознания, завуалированное морфологией"**.
* (Термин был предложен еще в середине прошлого века американским геологом и палеонтологом Дж. Дана. См.: D. С. Gilman. The life of James Dwight Dana. N. Y.- London, 1889.)
** (П. Тейяр де Шардэн. Феномен человека. М., "Прогресс", 1965, стр. 167.)
Тейар де Шардэн рисует эволюцию как ортогенетический процесс, вызываемый непостижимыми силами и ведущий к человеку. Благодаря человеку биосфера постепенно переходит в новое состояние - "разумную сферу", или ноосферу. Понятие ноосферы ввел в 1927 г. французский философ Е. Леруа; В. И. Вернадский в 1944 г. писал, что под влиянием возрастающей деятельности человека биосфера вступила в новую стадию своего развития - стадию ноосферы (см. главу 27).
Финалисты противопоставляют свои взгляды современной синтетической теории эволюции, сторонники которой будто бы строят эволюцию на случайности и не способны объяснить прогресс. В противовес этому финалисты прибегают к нематериальному фактору - "антислучайности" (см. главу 17).
Проблема эволюционного прогресса, получившая идеалистическое толкование у финалистов, широко и успешно разрабатывалась в Советском Союзе. А. Н. Северцов (1931), И. И. Шмальгаузен (1939) и другие биологи показали, что поступательное развитие живой природы нисколько не противоречит дарвинизму и служит блестящей иллюстрацией диалектико-материалистического мировоззрения (см. главы 17 и 19).
Выше говорилось преимущественно о достижениях в познании основных событий истории органического мира в рамках крупных эволюционных преобразований, именуемых обычно макроэволюцией. В изучении микроэволюции успехи палеонтологии гораздо скромнее. Усиление работ в этой области отвечает запросам современного эволюционного учения. Всестороннее изучение вида в пространстве и времени на массовом палеонтологическом материале при помощи ЭВМ и математических методов призвано также удовлетворить насущные потребности стратиграфии.
Палеомагнитные наблюдения, данные о палеотемпературах и другие новые материалы о климатических поясах Земли в прошлом открывают широкие перспективы в изучении населения Земли и эволюции биогеоценозов по природным зонам от архея до антропогена. Большие сдвиги происходят в палеонтологии в последнее время в результате разработки и применения новых методов препаровки и исследования микроструктуры и химического состава скелетных остатков. Методами электронной микроскопии и хроматографии в конхиолине раковин моллюсков устанавливается состав аминокислот, влияющих на микроструктуру. Это также открывает новые возможности для исследования филогении и систематики.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'