7.2.5. Эндоплазматический ретикулум (ЭР)
Одним из самых важных открытий, сделанных с помощью электронного микроскопа, было обнаружение сложной системы мембран, пронизывающей цитоплазму всех эукариотических клеток. Эта сеть мембран, получившая название эндоплазматический ретикулум (лат. reticulum-сеть), очень хорошо развита в клетке, но лежит за пределами разрешающей способности светового микроскопа. Сразу же было отмечено, что мембраны усеяны какими-то мелкими частицами, которые позже стали известны под названием "рибосом". Приблизительно в то же время методом дифференциального центрифугирования была получена клеточная фракция, способная осуществлять синтез белка. Изучение этой фракции с помощью электронного микроскопа показало, что она состоит из множества мелких мембранных мешочков (везикул), покрытых снаружи рибосомами. Эти мембранные мешочки были названы микросомами. Теперь мы знаем, что появление микросомной фракции - это результат процесса гомогенизации. Когда при гомогенизации ЭР распадается на мелкие фрагменты, края этих фрагментов смыкаются и образуются везикулы. В интактных клетках микросом нет.
На ультратонких срезах ЭР имеет вид множества парных параллельных линий (мембран), располагающихся в цитоплазме (рис. 7.4.-7.6). Однако иногда срез проходит так, что мы получаем возможность посмотреть как бы сквозь поверхность этих мембран, и тогда можно видеть, что в трех измерениях ЭР имеет не трубчатое, а пластинчатое строение. Модель трехмерной структуры ЭР изображена на рис. 7.16. ЭР состоит из уплощенных мембранных мешочков, называемых цистернами. Цистерны ЭР могут быть покрыты рибосомами, и тогда он называется шероховатым ЭР; если рибосомы отсутствуют, то его называют гладким ЭР (строение ближе к трубчатому). Функция обоих типов ЭР связана с синтезом и транспортом веществ.
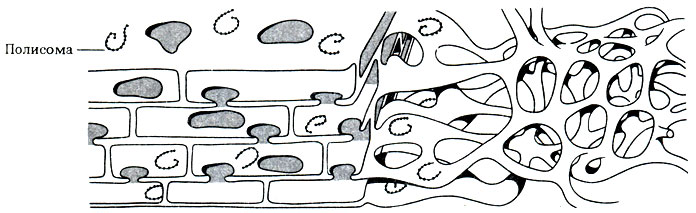
Рис. 7.16. Трехмерная модель эндоплазматического ретикулума
Функции шероховатого эндоплазматического ретикулума связаны с транспортом белков, синтезируемых рибосомами на его поверхности. Подробно белковый синтез описан в гл. 22. Здесь достаточно сказать, что растущая белковая молекула, т. е. цепь из аминокислот, или так называемая полипептидная цепь, остается присоединенной к рибосоме до тех пор, пока ее синтез не завершится. В начале синтеза белка первую часть растущей цепи может составлять "сигнальная последовательность", соответствующая по своей конфигурации специфическому рецептору на мембране ЭР и благодаря этому обеспечивающая связывание рибосомы с ЭР. Рецептор образует канал, по которому белок переходит в цистерны ЭР (рис. 7.17). Как только белок попадет внутрь, сигнальная последовательность отделяется от полипептидной цепи, и белок, свертываясь, приобретает в цистернах ЭР свою третичную структуру.
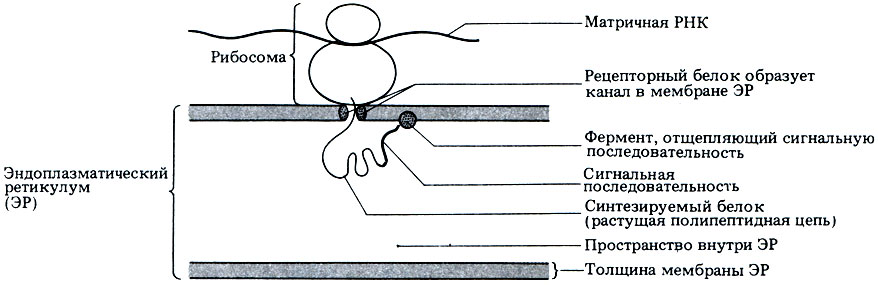
Рис. 7.17. Поступление новосинтезированного белка в эндоплазматический ретикулум
7.5. Значительную долю в сигнальной последовательности аминокислот составляют неполярные аминокислоты. Как вы могли бы это объяснить?
Транспортируясь затем по цистернам, белок обычно претерпевает на своем пути весьма существенные изменения. Он может, например, фосфорилироваться или превращаться в гликопротеин. Обычный путь для белка - это путь через шероховатый ЭР в аппарат Гольджи, откуда он либо выходит из клетки наружу (секретируется), либо поступает в другие органеллы той же клетки, например в лизосомы или откладывается в виде запасных гранул.
Белок, не имеющий сигнальной последовательности, синтезируется свободными рибосомами и выделяется в цитозоль для использования в этой же клетке.
Одной из главных функций гладкого ЭР является синтез липидов. Так, в эпителии кишечника гладкий ЭР синтезирует липиды из жирных кислот и глицерола, всасывающихся в кишечнике, а затем передает их в аппарат Гольджи для экспорта. Стероиды - это один из классов липидов, поэтому гладкий ЭР обильно представлен в тех клетках, которые секретируют стероидные гормоны, например в клетках коры надпочечников или в интерстициальных клетках семенников. В печени как шероховатый, так и гладкий ЭР участвуют в процессах детоксикации. В мышечных клетках присутствует особая, специализированная форма гладкого ЭР - так называемый саркоплазматический ретикулум (разд. 17.4).
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'