12.3.7. Продуктивность
Изучая продуктивность экосистем, мы имеем дело с потоком энергии, проходящим через ту или иную экосистему.
Энергия поступает в биотический компонент экосистемы первичных продуцентов. Скорость накопления энергии первичными продуцентами в форме органического вещества, которое может быть использовано в пищу, называется первичной продукцией. Это важный параметр, так как им определяется общий поток энергии через биотический компонент экосистемы, а значит, и количество (биомасса) живых организмов, которые могут существовать в экосистеме.
12.9. Какие группы организмов, помимо растений, принимают участие в создании первичной продукции экосистемы?
Как уже упоминалось в разд. 12.3.3, количество солнечного света, падающего на растения, различно. Оно зависит от географической широты и от степени развития растительного покрова и на Британских островах составляет (в энергетическом выражении) 1 ⋅ 106 кДж/м2 в год. Примерно 95-99% этого количества сразу отражается, поглощается с переходом в тепло или расходуется на испарение воды, и лишь 1-5% поглощается хлорофиллом и используется для синтеза органических молекул. Скорость, с которой растения накапливают химическую энергию, называется валовой первичной продуктивностью (ВПП). Примерно 20% этой энергии расходуется растениями на дыхание и фото дыхание. Скорость накопления органического вещества за вычетом этого расхода называется чистой первичной продуктивностью (ЧПП). Это энергия, которую могут использовать организмы следующих трофических уровней.
В примере, приведенном на рис. 12.12, ЧПП составляет 8000 кДж/м2 в год, т. е. 0,8% от полученной солнечной энергии (1% поглощенного излучения и 80% ВПП). В схеме указаны среднегодовые показатели, так как продуктивность летом выше, чем зимой.
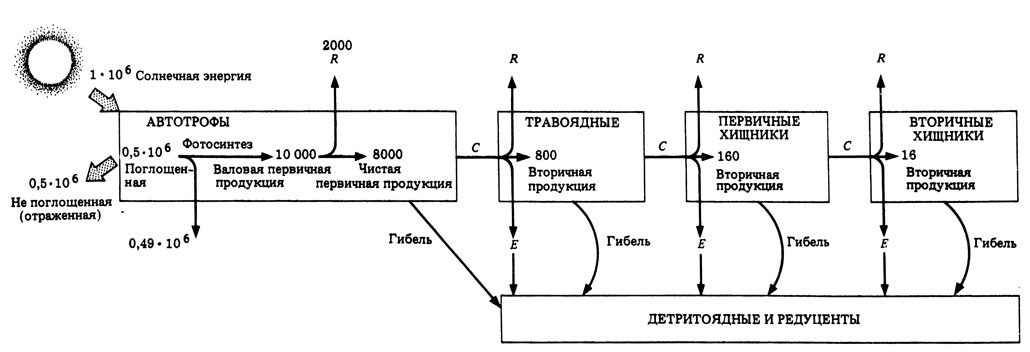
Рис. 12.12. Поток энергии через пастбищную пищевую цепь. Размерность цифр - кДж⋅м-2⋅год-1
При поедании одних организмов другими пища (вещество и энергия) переходит с одного трофического уровня на следующий. Непереваренная часть пищи выбрасывается. Животные, обладающие пищеварительным каналом, выделяют фекалии (экскременты); в них содержится некоторое количество энергии, так же как и в органических отходах метаболизма (экскретах), например мочевине.
Животные, как и растения, теряют часть энергии при дыхании. Энергия, оставшаяся после потерь, связанных с процессами дыхания, пищеварения и экскреции, идет на рост, поддержание жизнедеятельности и размножение.
Количество органического вещества, накопленного гетеротрофными организмами, называется вторичной продукцией (на любом трофическом уровне). Приведенное ниже словесное уравнение суммирует расход потребленной животным энергии:
Некоторые члены этого уравнения нетрудно определить на домашних или диких животных в лабораторных условиях. Показателем роста за какое-то время служит увеличение биомассы или же энергии, заключенной в теле. Фекалии и экскреты можно собрать, взвесить и вычесть их массу из общей массы потребленной пищи, чтобы найти массу усвоенной пищи, использованной для роста и других процессов.
На рис. 12.12 ясно показано, что в каждом звене пищевой цепи часть энергии теряется. Отсюда очевидно, что длина пищевой цепи лимитируется размерами этих потерь. Отношение количества световой энергии, поглощаемой растениями, к чистой первичной продукции очень велико. Последующие переносы энергии по меньшей мере в десять раз эффективнее, чем первоначальный перенос. Средняя эффективность переноса энергии от растений к травоядным животным составляет около 10%, а от животного к животному - около 20%. В целом травоядные менее эффективно усваивают пищу, чем хищники. Это связано с тем, что растения содержат большое количество целлюлозы, а часто и древесины (содержащей целлюлозу и лигнин), которые плохо перевариваются и поэтому не могут служить источником энергии для большинства травоядных.
Энергия, теряющаяся при дыхании, не передается другим организмам. Энергия же, заключенная в экскрементах и отходах метаболизма, передается детритофагам и редуцентам и, таким образом, не теряется для экосистемы. Детритные цепи начинаются с мертвых организмов и отмерших частей растений (опавших листьев, ветвей). Доля чистой первичной продукции, переходящей прямо в детрит и в пищевые цепи редуцентов, в разных системах неодинакова. В лесной экосистеме большая часть первичной продукции поступает в детритные цепи, а не в пастбищные. Поэтому лесная подстилка оказывается местом значительной активности консументов, хотя эти организмы здесь обычно незаметны. Однако в морских экосистемах и на интенсивно используемых пастбищах больше половины первичной продукции может поступать в пастбищную пищевую цепь.
12.10. На рис. 12.13 показаны потоки энергии, проходящие через небольшую часть луговой экосистемы (цифры в килоджоулях на 1 м2 в год).
а) Какова валовая первичная продукция злаков и разнотравья?
б) Какова эффективность фотосинтеза (т.е. преобразования поступающей солнечной энергии в общую продукцию?
в) Чему равна чистая продукция птиц, питающихся семенами, паукообразных и кузнечиков (по отдельности)?
г) Сколько энергии теряется при дыхании и выделении фекалий у полевых мышей?
д) Какие организмы являются продуцентами, первичными консументами и вторичными консументами?
ё) Какие организмы относятся к гетеротрофам?
ж) Какие еще могут быть пути для потоков энергии? (Назовите три.)
[Из Tribe М.А. et al. (1975), Ecological Principles, CUP.]
Детритные цепи сложнее и менее изучены, чем пастбищные. Но роль их с точки зрения потока энергии столь же велика, а часто и более значительна, чем роль пастбищных цепей.
Приведенные в этом разделе цифры - среднегодовые величины. Если экосистема стабильна и в ней не происходит увеличение общей биомассы, то биомасса в конце года будет такой же, как и вначале. Вся энергия, заключенная в первичной продукции, пройдет за это время через различные трофические уровни, а чистая продукция будет равна нулю. Обычно, однако, система изменяется. Например, молодой лес к концу вегетационного периода накапливает часть поступившей энергии в форме прироста биомассы. Продуктивность удобно выражать в среднегодовых показателях, так как при этом учитываются сезонные изменения продуктивности, если они есть. Например, первичная продукция больше в то время года, когда начинается массовая вегетация растений, а увеличение вторичной продукции происходит позднее.
Одна из причин, которые побуждают изучать поток энергии, проходящий через экосистему,- это возможность применения полученных знаний для удовлетворения потребности человека в пище и энергии. Научный анализ позволяет оценивать традиционные методы агротехники и вносить усовершенствования с целью повышения их эффективности. Поскольку на каждом трофическом уровне происходит потеря энергии, ясно, что для всеядных организмов (и в том числе для человека) наиболее эффективный способ извлечения энергии из экосистемы - потребление растительной пищи (табл. 12.3). Однако необходимо учитывать и другие факторы. Например, животный белок содержит больше незаменимых аминокислот, хотя некоторые бобовые культуры (например, соя) лишь незначительно уступают ему по своей ценности. Кроме того, растительный белок переваривается труднее, чем животный, так как, прежде чем извлечь его, необходимо разрушить жесткие клеточные стенки. Наконец, существует немало экосистем, где животные добывают пищу на большой территории и где было бы трудно выращивать культурные растения или собирать их урожай. Таковы, например, неплодородные земли, на которых пасутся овцы в Британии, северные олени в Шотландии и Скандинавии, канны в Восточной Африке.
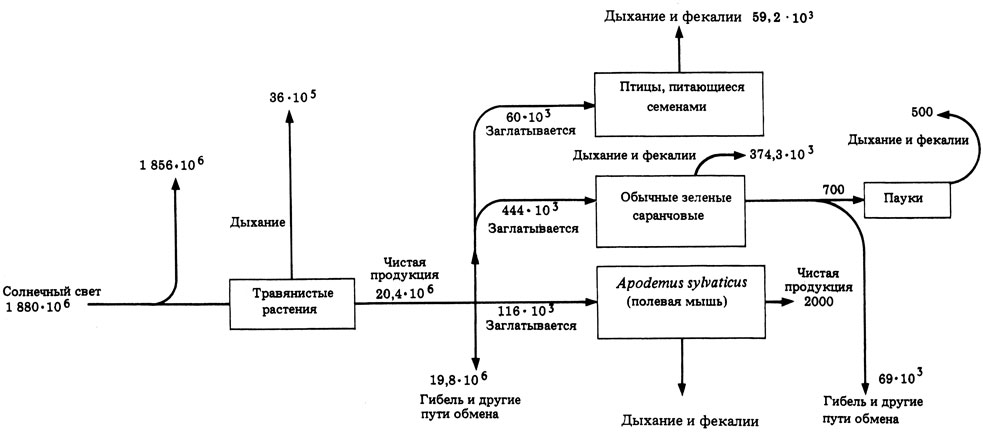
Рис. 12.13. Поток энергии через небольшую часть луговой экосистемы. Размерность цифр - кДж ⋅ м-2 ⋅ год-1. (Из M.R. Tribe et all (1974). Ecological principles, Basic Ecology Course 4, Cambridge University Press.)

Таблица 12.3. Продукция сельскохозяйственных пищевых цепей в Соединенном Королевстве*
* (Данные из Duckham, A.N. & Mansfield, G. В. (1970) Farming Systems of The World, Chatto and Windus.)
"Получение урожая" означает изъятие из экосистемы тех организмов или их частей, которые используются в пищу (или для иных целей). При этом желательно, чтобы экосистема производила пригодную для пищи продукцию наиболее эффективно. Это может быть достигнуто путем повышения урожайности культуры, уменьшения заболеваемости и помех со стороны других организмов или использования культуры, более приспособленной к условиям данной экосистемы.
Для повышения урожаев культурных растений могут применяться различные средства в зависимости от конкретных условий. Можно, например, вносить в почву удобрения, осуществлять искусственное орошение или, наоборот, удалять избыток воды через дренажную систему. Однако длительное применение искусственных удобрений может привести к ухудшению структуры почвы (разд. 12.4.1), а это ведет к уменьшению ее плодородия. Внесение удобрений и воды часто требует больших затрат энергии.
Уменьшение вреда от насекомых и птиц, а также от болезней достигается выведением генетически устойчивых сортов и применением химических средств, уничтожающих возбудителей болезней и вредителей. Однако такие средства нужно использовать осторожно. Применение стойких препаратов, неспособных быстро разлагаться в экосистеме, может иметь катастрофические последствия на других трофических уровнях (разд. 12.3.8). Частое применение эффективного пестицида может вызывать вспышки численности устойчивых к нему форм вредителей, с которыми труднее бороться. Такая ситуация возникла в Африке при попытках контролировать численность малярийного комара с помощью ДДТ.
Примером использования культур, более приспособленных к окружающей среде, служит разведение в условиях относительно высокой интенсивности света и температуры С4 - растений, таких, как сахарный тростник и кукуруза, а не С3 - растений. В этих условиях эффективность фотосинтеза и соответственно продуктивность С4 - растений выше, чем у С3 - растений (разд. 9.8.2). Следующий пример объясняет, почему в Восточной Африке может быть выгоднее использование диких копытных, чем разведение домашнего скота. Дикие копытные, например канна, имеют в течение года большую биомассу (82,2 - 117,5⋅105 кг/га по сравнению с 13,2 - 37,6⋅105 кг/га для домашних животных), так как они эффективнее переваривают пищу и быстрее усваивают питательные вещества, питаясь бедными по составу травами, что особенно важно в течение сухого сезона в саванне. Дикие копытные, вероятно, также более устойчивы к заболеваниям, встречающимся в районе их обитания.
Большую роль при разведении животных, достигающих половой зрелости через несколько лет, играет концепция максимального постоянного уровня добычи. Это наибольшая скорость изъятия особей из популяции без ущерба для ее дальнейшей продуктивности. Для определения этой величины необходимо иметь ясное представление о структуре и динамике популяции (разд. 12.6), особенно о рождаемости: ведь чем выше рождаемость, тем больше особей можно изымать из популяции. Знание структуры популяции позволяет установить, какие возрастные группы играют наибольшую роль в воспроизводстве стада, и соответственно отбирать для убоя или отлова животных других возрастных групп. В Англии, например, официально установлен минимальный размер ячеек для сетей, которыми можно пользоваться в промысловом рыболовстве. Таким образом, вылавливается более крупная и старая рыба, которая уже дала потомство, а не достигшие половой зрелости особи в сеть, как правило, не попадают. Вылов молоди привел бы к уменьшению воспроизводства рыбного стада в последующие годы и, таким образом, уменьшил бы размеры популяций в будущем.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'