12.6.6. Взаимодействия между популяциями
Редко изучают популяционную динамику только одного вида. Как мы уже видели, для понимания колебаний численности зимней пяденицы нужно также кое-что знать о ее паразитах. Существует ряд хорошо изученных типов взаимодействий между популяциями различных видов (межвидовые взаимодействия). На данном трофическом уровне это может быть межвидовая конкуренция, т.е. конкуренция между особями различных видов за доступные ресурсы, например пищу и пространство. Именно поэтому для экологии сообществ важно изучение ниш. Популяции, находящиеся на разных трофических уровнях, тоже могут взаимодействовать - возможны, например, отношения типа "хищник-жертва" и "хозяин-паразит". Существуют и другие типы взаимоотношений; иногда они трудноуловимы и сложны, как, скажем, некоторые симбиотические взаимоотношения, из которых оба партнера извлекают пользу.
Взаимодействуют между собой и особи одного вида (внутривидовые взаимодействия). Сюда относятся территориальные отношения и другие формы конкурентного поведения.
Для изучения межвидовых взаимодействий можно создавать в лаборатории простые и легко контролируемые ситуации, рассчитывая на то, что они окажутся подходящими моделями реальных ситуаций.
В качестве примера приведем простую и часто используемую модель взаимоотношений "хищник- жертва", реализуемую в экспериментах с двумя клещами, один из которых хищный (Typhlodromus), а другой травоядный (Eotetranychus). На рис. 12.39 показаны циклические колебания их численности. Циклы этих двух видов несколько не совпадают по фазе друг с другом.
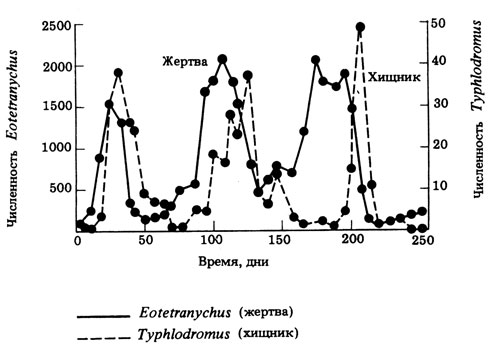
Рис. 12.39. Колебания численности популяций хищного клеща Typhlodromus и его жертвы - растительноядного клеща Eotetranychus-в лабораторных условиях. (М. К. Sands (1978) Problems in Ecology, Mills, Boon Ltd.)
Такие циклы можно объяснить следующим образом: возрастание численности жертвы ведет к увеличению популяции хищника, что в свою очередь вызывает резкое снижение численности жертвы, а за ним неизбежно следует уменьшение популяции хищника; цикл замыкается, когда уменьшение числа хищников позволяет снова вырасти числу жертв. Каждый цикл включает несколько поколений. Выводы, основанные на изучении таких лабораторных моделей, можно лишь с большой осторожностью переносить на отношения "хищник - жертва" в природе, где могут действовать и другие факторы. Однако регулярные циклические колебания численности действительно характерны для популяций некоторых травоядных млекопитающих. Например, у леммингов в Северной Америке и Скандинавии наблюдается четырехгодичный цикл. Поэтому также циклически колеблется и численность их врагов - песца и белой совы. Однако численность леммингов периодически сокращается, скорее всего из-за истощения кормовых ресурсов, а не из-за хищников. Время от времени, когда популяции скандинавских леммингов достигают исключительно высокой плотности, они массами мигрируют, и многие при этом гибнут в фиордах или тонут в реках. На циклические изменения популяций других травоядных тоже может в большей степени влиять наличие корма, нежели хищники. Фактором, вызывающим колебания численности, могут быть также болезни. Этот фактор зависит от плотности популяций, так как вероятность распространения эпизоотий возрастает с ее увеличением. Судя по некоторым данным, эпизоотии могут играть роль и в циклических колебаниях популяции американского зайца-беляка в северной Канаде. Здесь численность главного врага зайцев - канадской рыси - колеблется сходным образом, но со сдвигом по фазе. Зайцы составляют 80-90% пищевого рациона рыси.
Хищники, несомненно, играют важную роль в регулировании численности животных, хотя это, возможно, и не единственный фактор. На примере популяции оленей, обитающих на плато Кайбаб в Аризоне, можно судить о важности отношений "хищник - жертва" и о тех долговременных преимуществах, которые эти отношения доставляют популяции жертвы. В 1906 г. район плоскогорья был объявлен заповедником, и, для того чтобы защитить оленей от хищников (пумы, росомахи, койота) последних систематически истребляли в течение 30 лет. До 1906 г. популяция оставалась стабильной и состояла примерно из 4 тыс. особей, но впоследствии произошел "демографический взрыв" (рис. 12.40), и поддерживающая емкость среды, в которой по оценкам могла существовать 30-тысячная популяция оленей, оказалась недостаточной. В 1924 г. численность оленей достигла 100 тыс. особей, т.е. рост ее шел по J-образной кривой. Однако в результате истощения кормовых ресурсов вскоре начался голод, который вместе с болезнями привел к резкому падению численности популяции. Растительность была серьезно повреждена и уже не восстановилась до уровня 1906 г.; в результате поддерживающая емкость среды снизилась, и кормом теперь могло быть обеспечено только 10000 оленей.
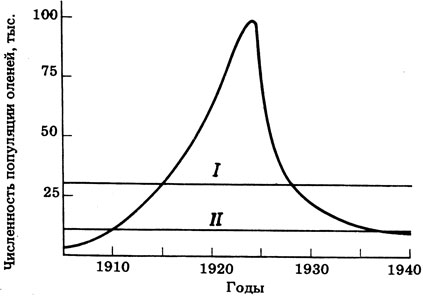
Рис. 12.40. Изменения численности популяций оленей на плато Кай баб, вызванные хищничеством. I - поддерживающая емкость среды в 1906 г; II - поддерживающая емкость среды в 1940 г.
Иногда наблюдаются циклические колебания численности паразита и хозяина. Фазы циклов, как и в примерах, описанных выше, не совпадают. Это особенно характерно для случаев паразитизма одних насекомых на других.
В экосистеме между популяциями возможна конкуренция за любой из доступных ресурсов, например за пищу, пространство, свет или убежища. Если два вида находятся на одном трофическом уровне, между ними весьма вероятна конкуренция за пищу. Со временем адаптивная эволюция одного или обоих видов может привести к тому, что каждый вид в пределах одного трофического уровня будет занимать отдельную нишу и конкуренция таким образом будет сведена к минимуму. В других случаях, когда конкурирующие виды занимают одинаковые или сильно перекрывающиеся ниши, может быть достигнуто равновесие, при котором ни один из видов не будет развиваться столь же успешно, как в отсутствие конкурента, или же численность одного из видов начнет снижаться и он будет полностью вытеснен. Последнее явление называют конкурентным исключением. Изучать это явление в популяциях диких организмов очень трудно, но был проведен ряд классических экспериментов с лабораторными популяциями. Первая такая работа принадлежит советскому биологу Гаузе (1934), который изучал конкурентные отношения между несколькими видами Paramecium. Некоторые результаты этой работы показаны на рис. 12.41.
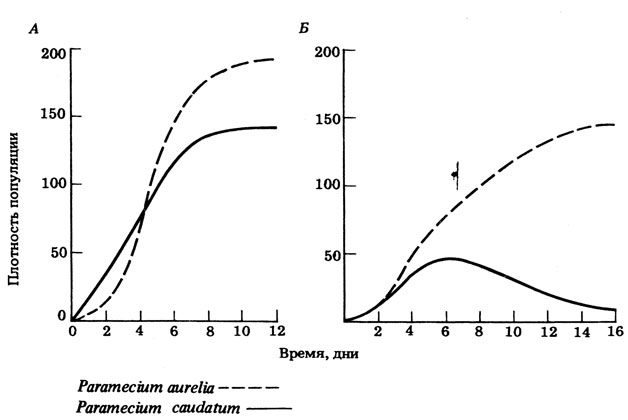
Рис. 12.41. Рост численности двух видов Paramecium. А. При выращивании видов отдельно. Б. При выращивании в одной культуре
12.20. Рассмотрите рис. 12.41.
а) Какой тип кривой роста популяции характерен для каждого из двух видов, когда они растут по отдельности?
б) За какие ресурсы идет конкуренция между двумя видами при их росте в одной культуре?
в) Какие факторы обеспечивают преимущество P. aurelia в конкуренции с P. caudatum!
При выращивании двух видов инфузорий в одной культуре вид P. aurelia оказывается более конкурентоспособным в захвате пищи, чем P. caudatum. Через пять дней численность P. caudatum начинает уменьшаться, спустя примерно 20 дней этот вид полностью исчезает, т.е. происходит его конкурентное исключение. P. aurelia достигает стационарной фазы роста позднее, чем при выращивании в отдельной культуре. Хотя этот вид и более конкурентоспособен, на него тоже отрицательно влияет конкуренция. Это помогает объяснить давление отбора, направленное на адаптацию конкурирующих видов к отдельным нишам. В природных условиях менее конкурентоспособный вид редко исчезает полностью - просто его численность уменьшается, а иногда может даже снова возрасти, прежде чем установится равновесное состояние.
Принцип конкурентного исключения (или принцип Гаузе) в дальнейшем был подтвержден другими экспериментами на животных. Конкурентное исключение наблюдается в популяциях растений, например в смешанных культурах видов ряски (Lemna); L. gibba способна вытеснять L. polyrrhiza.
Природные популяции изучать труднее, так как здесь одновременно взаимодействует большее число популяций, а параметры окружающей среды, такие, как температура, влажность и пищевые ресурсы, не поддаются контролю.
В популяциях растений одной из форм конкурентных взаимодействий, вызывающей большой интерес, является аллелопатия. Это частный случай более общего явления - выработки растениями и микроорганизмами разнообразных сложных органических молекул, влияющих на рост других живых организмов. Сюда входят антибиотики и ингибиторы роста, такие, как пенициллин, который вырабатывается грибом Penicillium и действует как антибиотик на грамположительные бактерии (см. разд. 2.2.2). Химическая конкуренция между микроорганизмами весьма сильна и взаимоотношения между ними очень сложны. Когда выделяемые вещества влияют на другие конкурирующие организмы того же трофического уровня, такое взаимодействие называют аллелопатией. Иногда при этом используются летучие ароматические вещества - вторичные метаболиты, выделяемые некоторыми растениями. Например, Мюллер в 1966 г. показал, что летучие терпены, выделяемые пахучими растениями чапараля (тип кустарниковых зарослей) в Калифорнии, адсорбируются на частицах почвы и задерживают прорастание или подавляют рост соседних растений. Фенольные соединения, которые выщелачиваются в почву из подстилки некоторых растений, оказывают такое же действие. Многие растения меловых лугов являются ароматическими и поэтому плохо поедаются травоядными. Это служит дополнительным фактором в сохранении богатого разнообразия травянистых растений. Недавнее изучение аллелопатических взаимодействий позволяет предположить, что они широко распространены как в пределах одного трофического уровня, так и между разными уровнями. К важным веществам, выделяемым растениями, относятся фенольные соединения, терпеноиды и алкалоиды. Интересно, что они часто небезразличны и для животных. Примером животного, извлекающего пользу из таких веществ, может служить бабочка-данаида Danaus plexippus. Ее гусеницы кормятся на ваточнике - растении, которое содержит вещества, действующие на позвоночных как сильный сердечный яд. Полагают, что ядовитость или неприятный вкус этого растения предохраняет его от поедания травоядными животными. Однако гусеницы данаид устойчивы к яду, они могут накапливать его в себе и сохранять после превращения во взрослую форму, и это в свою очередь защищает бабочек от поедания птицами. Яркая и характерная расцветка крыльев у данаиды служит предостережением для потенциальных врагов, т.е. действует как защитное приспособление (рис. 12.42). Побочным следствием оказалось то, что у некоторых других, уже неядовитых бабочек выработалась подражательная окраска, которая им тоже служит защитой от хищников. Этот случай показывает, какими сложными могут быть взаимодействия между видами. Растения нередко содержат токсичные химические вещества, к которым нечувствительны лишь немногие виды травоядных животных.

Рис. 12.42. Бабочка-данаида (Danaus plexippus)
Два других важных процесса, в которых происходит взаимодействие между цветковыми растениями и животными,- это распространение семян и опыление (разд. 20.2). Последнее служит хорошим примером распространенного явления, называемого коэволюцией, когда виды приспосабливаются друг к другу и со временем вырабатывают различные адаптации, обусловливающие взаимную зависимость и выгоду. В случае с ядовитыми растениями наиболее полезным и надежным приспособлением для устойчивых к яду животных может стать "обслуживание" этих растений, например опыление их, теми бабочками, гусеницы которых питаются данным растением.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'