14.4. Подъем воды по ксилеме
Ксилема цветковых растений состоит из двух типов структур, переносящих воду,- трахеид и сосудов. В разд. 8.2.1 мы уже говорили о том, как выглядят соответствующие клетки в световом микроскопе, а также на микрофотографиях, полученных с помощью сканирующего электронного микроскопа (рис. 8.11). Строение вторичной ксилемы (древесины) мы рассмотрим в разд. 21.6.6.
Ксилема вместе с флоэмой образует проводящую ткань высших растений. Эта ткань состоит из так называемых проводящих пучков, которые состоят из особых трубчатых структур. На рис. 14.15 показано, как устроены проводящие пучки и как они располагаются в первичном стебле у двудольных и однодольных растений.
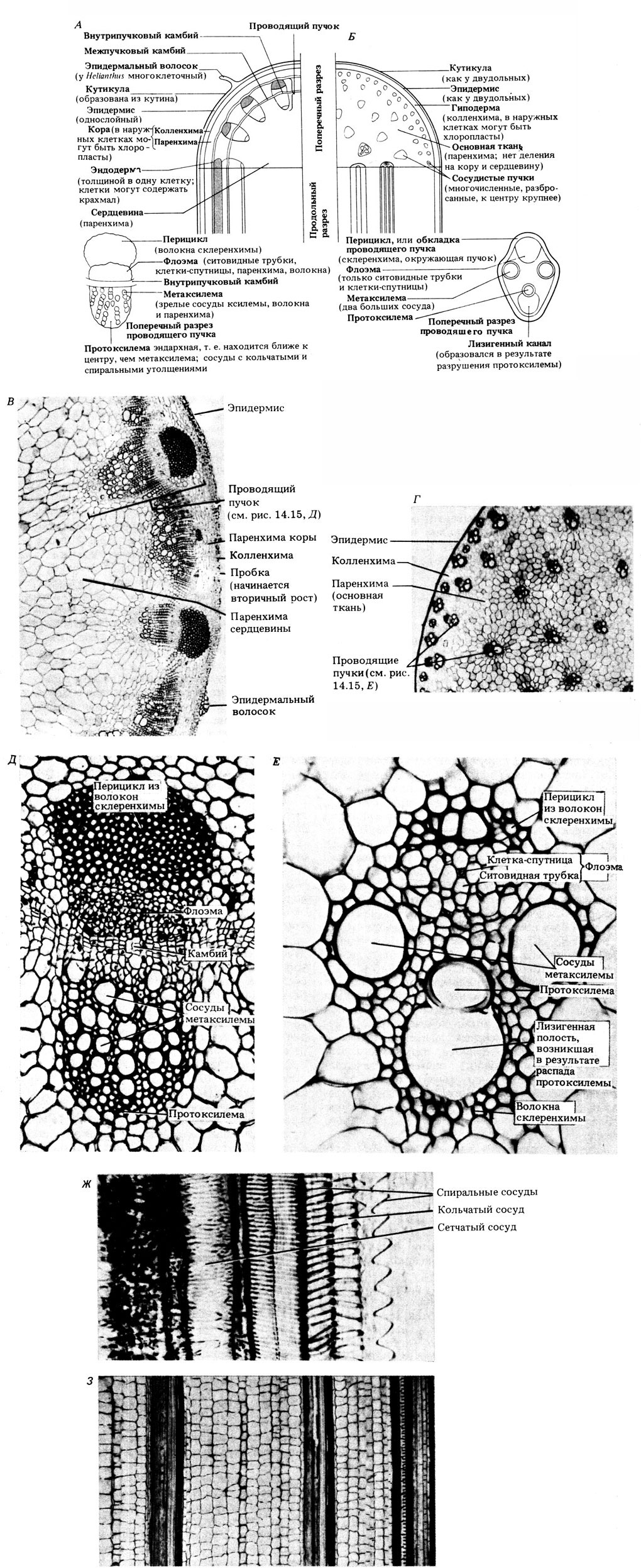
Рис. 14.15. А. Главные особенности анатомического строения стебля у типичного двудольного растения Helianthus annuus (подсолнечника). Б. Анатомическое строение стебля у типичного однодольного растения Zea mays (кукурузы). В. Часть поперечного среза стебля подсолнечника при малом увеличении. Г. Часть поперечного среза стебля Zea при малом увеличении. Д. Поперечный срез проводящего пучка из стебля подсолнечника при большом увеличении. Е. Поперечный срез проводящего пучка из стебля кукурузы при большом увеличении. Ж. Микрофотография продольного среза стебля подсолнечника. З. Микрофотография продольного среза стебля кукурузы
14.19. Резюмируйте в виде таблицы различия в строении первичного стебля у двудольных и однодольных растений.
14.20. Какова трехмерная форма следующих тканевых компонентов: а) эпидермиса; б) ксилемы; в) перицикла двудольных и г) сердцевины?
То, что вода может подниматься по ксилеме, очень легко продемонстрировать, погрузив нижний конец срезанного стебля в разбавленный раствор красителя, например эозина. Краситель поднимается по ксилеме и распространяется по всей сети листовых жилок. Если сделать тонкие срезы и просмотреть их в световом микроскопе, краситель будет обнаружен в ксилеме.
То, что ксилема проводит воду, лучше всего показывают опыты с "кольцеванием". Такие опыты проводили задолго до того, как стали применяться радиоактивные изотопы, позволяющие очень легко проследить путь вещества в живом организме. В одном из вариантов опыта вырезают кольцо коры с флоэмой. Если опыт не очень продолжителен, такое "кольцевание" не влияет на подъем воды по стеблю. Однако, если отслоить лоскут коры и вырезать ксилему, не повреждая лоскута коры, растение быстро завянет.
Любая теория, объясняющая передвижение воды по ксилеме, не может не учитывать следующие наблюдения:
1. Сосуды ксилемы-мертвые трубки с узким просветом, диаметр которого варьирует в пределах от 0,01 мм в "летней" древесине примерно до 0,2 мм в "весенней" древесине.
2. Большие количества воды переносятся относительно быстро: у высоких деревьев зарегистрирована скорость подъема воды до 8 м/ч, а у других растений она часто составляет около 1 м/ч.
3. Чтобы поднять воду по таким трубкам к вершине высокого дерева, необходимо давление порядка 4000 кПа. Самые высокие деревья - калифорнийские гигантские секвойи (хвойные, у которых нет сосудов и есть только трахеиды) и австралийские эвкалипты - бывают выше 100 м. Вода поднимается по тонким капиллярным трубкам благодаря высокому поверхностному натяжению под действием капиллярных сил; однако только за счет этих сил даже по самым тончайшим сосудам ксилемы вода не поднимется выше 3 м.
Все эти наблюдения удовлетворительно объясняет теория сцепления (когезии), или теория натяжения. Согласно этой теории, подъем воды от корней обусловлен испарением воды из клеток листа. Как мы уже говорили в разд. 14.3, испарение приводит к снижению водного потенциала клеток, примыкающих к ксилеме. Поэтому вода входит в эти клетки из ксилемного сока, у которого более высокий водный потенциал; при этом она проходит через влажные целлюлозные клеточные стенки сосудов ксилемы на концах жилок, как показано на рис. 14.7.
Сосуды ксилемы заполнены водой, и по мере того как вода выходит из сосудов, в столбе воды создается натяжения. Оно передается вниз по стеблю на всем пути от листа к корню благодаря сцеплению (когезии) молекул воды. Эти молекулы стремятся "прилипнуть" друг к другу, потому что они полярные и притягиваются друг к другу электрическими силами, а затем удерживаются вместе за счет водородных связей (разд. 5.1.2). Кроме того, они стремятся прилипнуть к стенкам сосудов под действием сил адгезии. Высокая когезия молекул воды означает, что для того, чтобы разорвать столб воды, требуется сравнительно большое растягивающее усилие; иными словами, столб воды обладает высокой прочностью на разрыв. Натяжение в сосудах ксилемы достигает такой силы, что может тянуть весь столб воды вверх, создавая массовый поток; при этом вода поступает в основание такого столба в корнях из соседних клеток корня. Необходимо, чтобы стенки сосудов ксилемы тоже обладали высокой прочностью и не вдавливались внутрь.
Такую прочность обеспечивают лигнин и целлюлоза. Данные о том, что содержимое сосудов ксилемы находится под воздействием большой растягивающей силы, были получены при измерении суточных изменений диаметра ствола у деревьев с помощью прибора, называемого дендрометром. Минимальные значения регистрировались в дневные часы, когда скорость транспирации максимальна. Крохотное сжатие отдельных сосудов ксилемы суммировалось и давало вполне измеримое уменьшение диаметра всего ствола.
Оценки прочности на разрыв для столба ксилемного сока варьируют в пределах примерно от 3000 до 30 000 кПа, при этом более низкие значения получены позднее. В листьях зарегистрирован водный потенциал порядка -4000 кПа, и прочность столба ксилемного сока, вероятно, достаточна, чтобы выдержать создающееся натяжение. Не исключено, конечно, что столб воды может иногда разрываться, особенно в сосудах большого диаметра.
Критики этой теории указывают на то, что любое нарушение непрерывности столба сока должно немедленно останавливать весь поток, так как сосуд должен заполняться воздухом и парами воды (явление кавитации). Кавитацию может вызвать сильное сотрясение, сгибание ствола или недостаток воды. Хорошо известно, что на протяжении лета содержание воды в стволе дерева постепенно уменьшается, а древесина заполняется воздухом. Это используется в лесной промышленности, потому что такое дерево обладает лучшей плавучестью. Однако разрыв водного столба в части сосудов не очень сильно влияет на скорость переноса воды. Это можно объяснить тем, что вода переходит из одного сосуда в другой или же обходит воздушную пробку, передвигаясь по соседним клеткам паренхимы и их стенкам. Кроме того, согласно расчетам, для поддержания наблюдаемой скорости потока вполне достаточно, чтобы в каждый момент времени функционировала хотя бы небольшая часть сосудов. У некоторых деревьев и кустарников вода передвигается только по самому молодому наружному слою древесины, который называют заболонью. У дуба и ясеня, например, вода движется главным образом по сосудам текущего года, а остальная часть заболони выполняет функцию водного резерва. В течение вегетационного сезона все время прибавляются новые и новые сосуды, но больше всего их образуется в начале сезона, когда скорость потока гораздо выше.
Вторая сила, которая участвует в передвижении воды по ксилеме,- это корневое давление. Его можно обнаружить и измерить в тот момент, когда отрезают крону, а штамб с корнями продолжает выделять сок из сосудов ксилемы. Этот процесс эксудации подавляется цианидом и другими ингибиторами дыхания и прекращается при недостатке кислорода или понижении температуры. Для работы такого механизма, по-видимому, нужна активная секреция в ксилемный сок солей и других водорастворимых веществ, снижающих водный потенциал. Затем в ксилему поступает вода за счет осмоса из соседних клеток корня.
Одного положительного гидростатического давления около 100-200 кПа (в исключительных случаях до 800 кПа), создаваемого за счет корневого давления, обычно недостаточно, чтобы обеспечить передвижение воды вверх по ксилеме, но его вклад у многих растений несомненен. У медленно транспирирующих травянистых форм этого давления, однако, вполне хватает для того, чтобы вызвать гуттацию. Гуттация - это выведение воды в виде капель жидкости на поверхности растения (тогда как при транспирации вода выходит в виде пара). Все условия, уменьшающие транспирацию, т. е. слабая освещенность, высокая влажность и т.п., способствуют гуттации. Она весьма обычна у многих растений влажных тропических лесов и часто наблюдается на кончиках листьев молодых проростков.
14.21. Перечислите те свойства ксилемы, благодаря которым она обеспечивает транспорт воды и растворенных в ней веществ на большие расстояния.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'