Глава 11
11.1. Те вещества, при гидролизе которых выделяется больше энергии, чем при гидролизе АТФ, способны переносить свои фосфатные группы на АДФ с образованием АТФ; а те, при гидролизе которых выделяется меньше энергии, чем при гидролизе АТФ, будут присоединять фосфатные группы от АТФ.
11.2. АТФ можно сравнить с аккумулятором в том смысле, что его производство требует энергии и он служит удобным кратковременным накопителем энергии. АТФ вырабатывается в процессе дыхания и может перемещаться в любую часть клетки, где необходима энергия, "разряжаться" (превращаясь в АДФ), а затем при дыхании вновь "заряжаться" (когда АДФ снова превращается в АТФ).
11.3. Дегидрогенирование.
11.4. Дегидрогеназы (см. табл. 6.3).
11.5. а) Фосфорилирование.
б) Изомеразы.
в) (1) Дегидрогенирование (или окисление).
(2) Фосфорилирование.
г) Один из витаминов группы В.
11.6. Для быстрой диффузии промежуточных продуктов из цитоплазмы в митохондрии и обратно.
11.7.
| Поступающие: | Выходящие: |
|---|---|
| Пировиноградная кислота | |
| Кислород | Двуокись углерода |
| Восстановленный переносчик водорода | Окисленный переносчик водорода |
| АДФ | АТФ |
| Фосфат | Вода |
11.8. Обеспечивает увеличение поверхности, всасывающей продукты переваривания пищи.
11.9. С повышением концентрации растворенных молекул будет повышаться осмотическое давление плазмы, а это может повлиять на многие физиологические процессы.
11.10. 1) В клетке пигмент отделен от более изменчивой химической среды плазмы крови.
2) Заключение пигмента внутрь клеток уменьшает вязкость крови и тем самым облегчает работу сердца.
11.11. Вначале при соприкосновении крови с водой градиент концентрации .кислорода между ними будет велик. Однако по мере дальнейшего параллельного течения обеих жидкостей градиент будет уменьшаться до тех пор, пока относительное насыщение кислородом крови и воды не станет одинаковым. В результате насыщение крови будет значительно ниже максимальной точки и поэтому будет недоста-точным (рис. 11.11 отв.).
11.12. Потому что часть объема легочной вентиляции составляет мертвое пространство.
11.13. У мелких животных велико отношение поверхности тела к его объему, что увеличивает потерю тепла; поэтому им требуется больше кислорода для поддержания постоянной температуры тела.
11.14. Для этого нужно сравнить потребление ими кислорода (в единицу времени), приходящееся на 1 г массы тела.
11.15.
| ПДК = | СО2 | = | 102 | = 0,70. |
| О2 | 145 |
11.16.
| ПДК = | СО2 | = | 2 | = ∞ |
| О2 | 0 |
11.17. Потому что на дыхание у человека обычно расходуются углеводы и жиры.
11.18. а) Частота дыхания равна около 17 в 1 мин.
б) Объем воздуха, обмениваемого за один вздох, равен 450 мл (в среднем).
в) Легочная вентиляция составляет 17 - 450 мл = 7,65 л в 1 мин.
г) Потребление кислорода выражено наклоном линии АВ. Таким образом, потребление кислорода равно 1500 мл за 4 мин, или 375 мл за 1 мин.
11.19.(а)

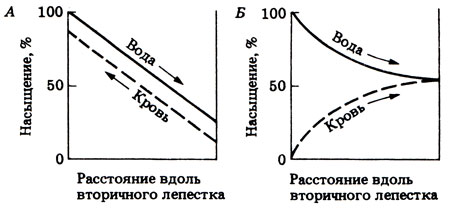
Рис. 11.11 (отв.). А. Противоток воды и крови. 6. Параллельный ток воды и крови
б) Перечень черт сходства между фотосинтезом и аэробным дыханием
Оба процесса приводят к преобразованию энергии.
Для обоих необходим механизм, обеспечивающий обмен двуокиси углерода и кислорода.
Для протекания обоих процессов у эукариот необходимы специальные органеллы, а именно митохондрии - для дыхания и хлоропласты - для фотосинтеза; митохондрии и хлоропласты сходны с прокариотическими организмами тем, что обладают кольцевой ДНК и прокариотическим типом белоксинтезирующей системы. Световые реакции фотосинтеза сходны с процессами клеточного дыхания в следующих отношениях:
1) в обоих случаях происходит фосфорилирование (т.е. синтез АТФ из АДФ и Фн);
2) те и другие процессы связаны с потоком электронов вдоль цепи переносчиков электронов;
3) для сопряжения переноса электронов с фосфорилирова- нием нужна определенная организация системы переносчиков в мембранах; в митохондриях такими мембранами служат кристы, в хлоропластах-тилакоиды.
Оба процесса связаны с циклическими цепями реакций, которые протекают в окружающем мембрану растворе (с циклом Кребса при дыхании, с циклом Кальвина при фотосинтезе). Часть ферментов гликолиза является общей для обоих процессов.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'