Глава 14
14.1. а) В растворе Б. б) Раствор А. в) Из Б в А. г) В растворе А. д) (2) -1000 кПа. Если это вас смущает, вспомните, что -1000 ближе к нулю, чем -2000. Нуль - это максимальное, наивысшее значение ???.
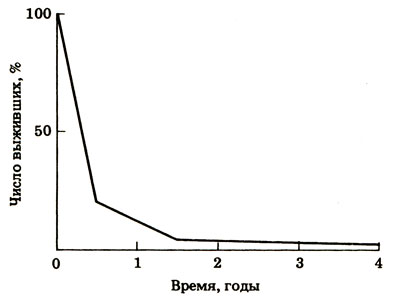
Рис. 12.18 (отв.). График, показывающий дорепродуктивную смертность среди нерки
е) В растворе Б. ж) Чем выше ОД раствора, тем ниже ψ (практически - ОД = ψ для раствора при нормальном атмосферном давлении).
14.2. Гипертоническим раствором. Вспомните, что клеточная стенка легко проницаема для растворов.
14.3. Нулевое. Протопласт не оказывает давления на клеточную стенку.
14.4. У прокариот (бактерий и сине-зеленых водорослей) и грибов. Хотя у прокариот нет вакуолей, их клетки обладают такими же осмотическими свойствами, что и клетки растений и грибов.
14.5. В начале ψклетки = ТД - ОД, а ψраствора = - ОД = = -1200 кПа; ТД = 0, так как клетка находится в состоянии начинающегося плазмолиза; ОД = 2000 кПа; таким образом, ψклетки = 0 кПа - 2000 кПа = -2000 кПа. Вначале у раствора выше, чем у клетки. Поэтому вода за счет осмоса пойдет из раствора в клетку. Единственный член уравнения, который существенно изменяется при переходе воды в клетку- это ТД. По мере поступления воды ТД, а следовательно и у, возрастает до тех пор, пока оно не станет препятствовать дальнейшему входу воды в клетку. В этот момент у клетки становится равным ψ раствора, и клетка становится тургесцентной.
Поэтому в состоянии равновесия ψклетки = - 1200 кПа. ψ = ТД - ОД, отсюда ТД = ψ + ОД = - 1200 кПа + + 2000 кПа = 800 кПа.
14.6. ОДклетки = 1100 кПа; ψ чистой воды = 0; В состоянии равновесия ψклетки = 0. ТД = ψ + ОД = 0 кПа + 1100 кПа = 1100 кПа.
14.7. а) ψклетки во время переноса = ψ чистой воды = 0, а ψ раствора сахарозы = - 800 кПа; отсюда разность ψ между содержимым клетки и наружным раствором равна 800 кПа.
б) Вода будет выходить из клетки (от более высокого у к более низкому).
в) ТД должно уменьшиться.
14.8. а) Клетка Б. б) Из клетки Б в клетку А.
в) В состоянии равновесия клетки должны иметь одинаковый ψ, он будет равен среднему от величин двух исходных ψ, т.е. -1000 кПа.
г) Клетка А в состоянии равновесия:
ТД = ψ + ОД = - 1000 кПа + 2000 кПа = 1000 кПа
Клетка Б в состоянии равновесия:
ТД = ψ + ОД = - 1000 кПа + 1400 кПа = 400 кПа
14.9. 1060 кПа. Для величин, занимающих промежуточное положение между значениями, приведенными в табл. 14.4, постройте график зависимости осмотического давления от молярной концентрации раствора сахарозы.
14.10. Усредненное ОД в клетках свеклы будет составлять около 1400 кПа.
14.11. ψ в клетках свеклы составит примерно - 940 кПа.
14.12. Можно получить более точный результат, взяв среднюю величину из двух или более повторностей. Некоторое представление о размерах вариаций, которых можно ожидать в этом случае, дают цифры, приведенные в табл. 14.6.
14.13. Чтобы предотвратить испарение воды и связанное с ним повышение концентрации растворов сахарозы, а также возможное высыхание полосок ткани.
14.14.
ТД = ψ + ОД = - 950 кПа + 1400 кПа = 450 кПа
Обратите внимание на то, что у разных корнеплодов свеклы могут быть разные значения ОД и ψ.
14.15. а) Клетки интактного стебелька тургесцентны, поэтому их стенки стремятся растянуться под действием тургорного давления. Толстые стенки эпидермальных клеток менее растяжимы, чем тонкие стенки клеток коры, и таким образом препятствуют расширению этих клеток. Клетки коры сжаты. Разрез эпидермиса снимает такое ограничение, все клетки коры слегка расширяются, общий объем коры увеличивается, и это приводит к тому, что полоски закручиваются наружу.
б) Дистиллированная вода имеет более высокий водный потенциал, чем клетки стебелька. Поэтому вода поступает в ткань за счет осмоса, еще больше растягивая клетки коры и увеличивая изгиб.
в) Концентрированный раствор сахарозы имеет более низкий водный потенциал, чем клетки стебелька. Поэтому вода в результате осмоса выходит из ткани, клетки коры сжимаются сильнее, чем клетки эпидермиса, и ткань изгибается внутрь.
г) Разбавленный раствор сахарозы должен иметь точно такой же водный потенциал, что и клетки стебелька; поэтому-то и не происходит чистого поступления воды или чистого выхода ее из раствора или из ткани.
д) Водный потенциал. План такого эксперимента может быть следующим.
Приготовьте ряд разбавленных растворов сахарозы, начиная с 1 М раствора и кончая дистиллированной водой (например: дистиллированная вода - 0,2 М- 0,4 М-0,6 М-0,8 М-1,0 М). Типичный изгиб свежесрезанных стебельков одуванчика следует зарегистрировать, зарисовав их; затем в каждый из растворов, разлитый по отдельным подписанным чашкам Петри, помещают по две полоски стебелька (лучше взять по две полоски, чтобы определить средний результат). После того как будет достигнуто равновесие (примерно через 30 минут), тщательно зарегистрируйте изгиб стебельков (например, зарисовав их). Тот раствор, в котором не произойдет никаких изменений, будет иметь тот же самый у, что и усредненная клетка стебелька в момент его среза.
14.16. План двух подходящих экспериментов может быть следующим.
Влияние температуры. Вырежьте кубики из свежего корнеплода свеклы, промойте их, чтобы удалить красный пигмент из поврежденных клеток, и поместите в стаканчики с водой разной температуры, скажем, в пределах от 20 до 100°С. Появление красного пигмента в воде будет указывать на нарушение избирательной проницаемости тонопласта (вакуолярной мембраны) и плазматической мембраны, приводящее к диффузии пигмента из клеточного сока в воду. Время, необходимое для появления определенного количества пигмента, позволит судить о скорости разрушения структуры мембран. Интенсивность окрашивания можно измерять на колориметре или же оценивать визуально.
Влияние этанола. Методика та же самая, только вместо изменения температуры используют ряд концентраций этанола.
14.17. а) Листья имеют множество устьиц, служащих для газообмена, и нет почти никаких препятствий для передвижения водяных паров через эти поры. б) У листьев большая поверхность (для того чтобы улавливать солнечный свет и для газообмена). Чем больше поверхность, тем значительнее потеря воды в результате транспирации.
14.18. Чем выше поднимается солнце, тем больше интенсивность света; она достигает максимума в полдень, когда солнце находится в верхней точке своего пути. Точно так же возрастает и температура воздуха, но с некоторым запаздыванием - примерно на два часа позже (главным образом потому, что сначала нагревается почва, а затем она излучает тепло в воздух). Первоначальное ускорение транспирации между 3 и 6 часами утра, еще до того, как повысится температура воздуха, обусловлено открытием устьиц на свету. С 6 часов утра нарастающая скорость транспирации тесно коррелирует с температурой (причины этого объясняются в тексте). Она слабо коррелирует с интенсивностью света, так как устьица в это время открыты полностью, и всякое дальнейшее повышение освещенности не оказывает никакого действия.
После полудня интенсивность света убывает, так как солнце начинает снижаться. Понижается и температура, но с такой же задержкой примерно в два часа. Транспирация замедляется и в результате понижения температуры, и в результате уменьшения освещенности, но она гораздо больше коррелирует со светом, ослабление которого приводит к постепенному закрыванию устьиц. Примерно к 19.30 наступает темнота, и устьица, вероятно, уже закрыты. Вся остальная транспирация, по-видимому, происходит через кутикулу, и на нее по-прежнему влияет температура.
14.19. См. табл. 14.19 (отв.).

Таблица 14.19 (отв.)
14.20. а) Тонкостенный полый цилиндр.
б) Массивный стержень (цилиндр), обеспечивающий опору.
в) Массивный стержень (цилиндр), обеспечивающий опору.
г) Массивный цилиндр.
14.21. 1) Ксилема состоит из длинных трубок, образованных в результате слияния соседних клеток и разрушения поперечных стенок между ними.
2) В трубках нет живого содержимого, отсюда меньшее сопротивление потоку.
3) Трубки обладают высокой прочностью, поэтому они не спадаются.
4) Тонкие трубки необходимы для того, чтобы водяной столб в них не разрывался.
14.22. См. табл. 14.22 (отв.).

Таблица 14.22 (отв.)
14.23. Почвенный раствор > клетка корневого волоска > клетка 3 > клетка 2 > клетка 1 > ксилемный сок.
14.24. а) При обеих температурах происходит быстрое начальное поглощение К+ (в первые 10-20 мин). Через 20 мин при 25°С продолжается непрерывное постепенное поглощение К+ , а при 0°С никакого дальнейшего поглощения не происходит. Поглощение при 25°С ингибируется KCN.
б) Есть две ясно различимые фазы поглощения. Во время начальной быстрой фазы К+ диффундирует в свободные пространства корня. Он проходит через клеточные стенки зоны корневых волосков, которые обращены к раствору, и диффундирует через апопласт (возможно, проникая в межклеточные пространства там, где в них находится раствор). Эксперименты показывают, что эта фаза мало зависит от температуры и, следовательно, не связана с обменом веществ (реакции, контролируемые ферментами, протекали бы при 25°С быстрее, чем при 0°С). Значит, эта первая фаза - пассивный процесс.
Вторая фаза зависит от температуры и не осуществлятся при 0°С, когда интенсивность метаболизма очень низка. Это позволяет предполагать, что поглощение требует затраты энергии, а его ингибирование KCN указывает на то, что оно зависит от дыхания. Таким образом, вторая фаза представляет собой активный транспорт через клеточную мембрану внутрь клетки.
14.25. Увеличение интенсивности дыхания сопровождается усилением поглощения КСl. Если КСl достаточно, он, по-видимому, поглощается путем активного транспорта за счет энергии, которую доставляет дыхание.
14.26. KCN, ингибируя дыхание, тем самым подавляет и активный транспорт КСl в высечки из моркови.
14.27. Значительная часть фосфата внутри корня находилась в свободном пространстве и могла поэтому диффундировать обратно в окружающую воду, обращая вспять пассивное поглощение.
14.28. Нет. Эндодерма - это барьер для передвижения воды и растворенных в ней веществ по апопластному пути (см. разд. 14.5.2).
14.29. Радиоавтография выявляет локализацию иона в тонких срезах. Ингибируйте у одного из растений активный транспорт (например, с помощью KCN или низкой температуры), а другое растение возьмите в качестве контроля. Пусть теперь оба растения поглощают радиоактивный ион. У растения с подавленным активным транспортом ионы будут передвигаться только пассивно через клеточные стенки. Радиоавтография должна показать, что радиоактивный ион почти не проникнет здесь дальше эндодермы, тогда как в контроле должно быть видно гораздо большее проникновение ионов в ткань за эндодермой.
14.30. а) См. табл. 14.30 (отв.).
Черты сходства в экспорте 14С из верхнего и из нижнего листа. Оба листа экспортируют одинаковую долю ассимилятов (сравните результаты для "листа, обработанного 14С"), и одинаковая доля переходит в другие листья. Главным местом назначения ассимилятов в обоих случаях является боб.
Различия в экспорте 14С из верхнего и из нижнего листа. На экспорт ассимилятов из того или иного листа влияет расположение этого листа на стебле. Верхний лист экспортирует большую долю своих ассимилятов в боб и апикальную зону стебля, чем нижний лист, и меньшую долю - в корни. б) Когда боб уже образовался, он становится существенным потребителем ассимилятов, особенно из тех листьев, которые находятся в непосредственной близости к нему. У взрослого растения есть еще два важных места потребления ассимилятов, а именно апикальная зона стебля и корни. Нижние листья экспортируют ассимиляты главным образом в корни, а верхние - в основном к верхушке стебля.
14.31. 0,72 с. Этот ответ получается таким образом:
100 см = 1000 мм = 1 000000 мкм = 106 мкм; следовательно, сахароза движется со скоростью 106 мкм/ч
= 1 мкм за 1/106 ч = 200 мкм за 200/106 ч = 2/104 ч = (2 ⋅ 3600)/104 с = 2 ⋅ 0,36 с = 0,72 с.
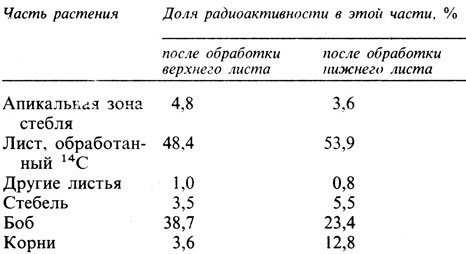
Таблица 14.30 (отв.)
14.32. Через 2500 ситовидных пластинок на каждый метр:
1 м = 106 мкм; 400 мкм = 4⋅102 мкм, 106/(4⋅102)= 104/4 = 2500.
14.33. Благодаря высокому давлению оксигенированная кровь может поступать в ткани достаточно быстро, чтобы удовлетворить метаболические потребности организма. Кроме того, на высоком гидростатическом давлении крови основана нормальная работа таких органов, как почки и кровеносные капилляры.
14.34. В большом круге кровообращения кровь поступает в капилляры под значительно более высоким давлением, а относительно низкое давление крови в легочной артерии предотвращает разрыв нежных легочных капилляров.
14.35. Благодаря расширению кровеносных сосудов в поврежденном участке улучшается его снабжение оксигенированной кровью и питательными веществами и быстрее идет процесс заживления. Повышение общего кровяного давления подготавливает организм животного к более быстрому и эффективному ответу на любой последующий стресс.
14.36. Перед стартом. Ожидание старта вызывает выброс адреналина. Под действием адреналина сужаются все кровеносные сосуды, кроме тех, которые снабжают кровью жизненно важные органы, и как следствие - повышается кровяное давление. Повышается частота сокращений сердца. Из селезенки в общую циркуляцию выбрасывается дополнительное количество крови. Во время бега. В это время повышается метаболическая активность, особенно в скелетных мышцах. Углекислота, образующаяся в мышцах в повышенных количествах, вызывает местное расширение кровеносных сосудов. В то же время повышение концентрации СО2 в системной крови регистрируется хеморецепторами аорты и каротидных телец, и сигналы от них стимулируют сосудодвигательный центр. Стимуляция этого центра приводит к сужению сосудов, повышению кровяного давления и ускорению кровотока. Увеличивается также частота и сила сердечных сокращений, и желудочки сердца полнее освобождаются от крови. К концу пробега в мышцах преобладает анаэробное дыхание и накапливается молочная кислота (разд. 11.3.8). Сильные сокращения мышц приводят к ритмическому сдавливанию вен, что способствует ускорению венозного возврата крови в сердце. Восстановительный период. Полностью ликвидируется кислородная задолженность, и молочная кислота удаляется из крови; уменьшается активность тканей и снижается содержание СО2. В результате этого частота сердечных сокращений и кровяное давление возвращаются к норме.
14.37. В активно дышащих тканях существует высокое парциальное давление СО2, которое приводит к снижению рН и смещению кривой диссоциации оксигемоглобина вправо. Анализ рис. 14.34 показывает, что в этих условиях облегчается переход кислорода из крови в ткани, где он может быть использован для выработки энергии в процессе дыхания. При низком парциальном давлении О2 гемоглобин связывает его слабо, но, будучи насыщен кислородом, легко его отдает.
Высокая метаболическая активность приводит также к повышению температуры в данном участке тела, а это ведет к уменьшению сродства гемоглобина к О2 и усиленной диссоциации оксигемоглобина. В результате кривая диссоциации тоже сдвигается вправо, и это имеет физиологический смысл, так как в активные участки поступает больше кислорода из крови. Кривые диссоциации оксигемоглобина у разных видов не совсем одинаковы; например, у мелких млекопитающих они сдвинуты вправо по сравнению с человеком. У мелких млекопитающих метаболическая активность намного выше, чем у человека, поэтому кислород должен у них освобождаться намного быстрее.
14.38. Сдвиг кривой диссоциации у плода по отношению к ее положению у матери означает, что кровь плода обладает большим сродством к О2, чем кровь матери, и это вполне естественно, поскольку плод должен получать весь кислород через плаценту из материнской крови. Поэтому при любом парциальном давлении кислорода кровь плода будет поглощать его из материнской крови и всегда будет сильнее насыщена кислородом, чем материнская кровь.
14.39. У южноамериканских лам кровь отличается высоким сродством к кислороду и способна связывать его при низком парциальном давлении, существующем на больших высотах. Это еще один яркий пример физиологической адаптации.
14.40. 1. Карбоксигемоглобин попадает в легкие, насыщается кислородом и превращается в оксигемоглобин.
2. Оксигемоглобин, будучи более слабым основанием, чем карбоксигемоглобин, освобождает ионы Н+ .
3. Ионы Н+ присоединяются в эритроцитах к ионам гидрокарбоната с образованием угольной кислоты.
4. Угольная кислота диссоциирует на СО2 и воду.
5. В результате потери ионов гидрокарбоната эритроцитами в эритроциты из плазмы диффундируют новые ионы гидрокарбоната.
6. Из гидрокарбоната в эритроцитах образуются новые молекулы угольной кислоты, которые опять диссоциируют на СО2 и воду.
7. СО2 диффундирует из эритроцитов и в конечном счете выводится из организма через легкие.
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'