Глава 15
15.1. Передвижение прежде всего обусловлено необходимостью искать пищу (и тесно связано с развитием нервной системы). Зеленые растения - автотрофные организмы, они способны сами синтезировать нужные им органические вещества, так что им нет надобности искать органическую пищу.
15.2. См. табл. 15.2 (отв.).
| Примеры | Преимущества |
|---|---|
| Побеги и колеоптили обладают положительным фототропизмом, а корни - отрицательным | Листья оказываются на свету, который служит источником энергии для фотосинтеза. Обнаженные корни растут в сторону почвы или иного подходящего субстрата |
| Побеги и колекоптили проявляют отрицательный геотропизм, а корни - положительный | Побеги и проростки растут вверх, т.е. к свету, а корни врастают глубже в почву |
| Корневищам, усам и т.п. свойствен диагеотропизм; листьям двудольных - тоже | Это помогает растениям заселять новые участки почвы; листья растут горизонтально и получают максимум света |
| Боковые корни и разветвления стебля проявляют плагиогеотропизм | Корни используют больший объем почвы, и их расположение усиливает их опорную функцию; листья на ветвях занимают больше пространства для использования света |
| Гифы грибов и пыльцевые трубки обнаруживают положительный хемотропизм | Гифы растут по направлению к пище, а пыльцевые трубки-к семязачатку (где происходит оплодотворение) |
| Корням и пыльцевым трубкам свойствен положительный гидротропизм | Вода нужна для всех жизненных процессов |
| Усики лиан проявляют положительный гаптотропизм; чувствительные волоски росянки - тоже | Это позволяет уськам выполнять опорную функцию, а волоскам - улавливать насекомых |
| Пыльцевым трубкам свойствен отрицательный аэротропизм | Трубка сразу же начинает расти в сторону ткани пестика (дальше от воздушной среды) |
15.3. Рост мог бы ингибироваться на стороне контакта или же стимулироваться на противоположной стороне (или могло бы происходить то и другое). Фактически рост замедляется (т.е. клетки меньше удлиняются) в зоне контакта, а рост на противоположной стороне ускоряется в 40-200 раз.
15.4. Возможны самые разные методики. Простейший эксперимент показан на рис. 15.4 (отв.).
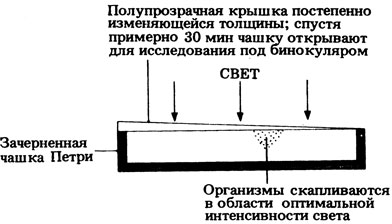
Рис. 15.4 (отв.) Опыт, позволяющий выяснить, какую интенсивность света предпочитает эвглена или хламидомонада
15.5. a) Spirogyra (или любая другая нитчатая зеленая водоросль).
б) Бактерии - аэробы и проявляют положительный аэротаксис. Поэтому они движутся в сторону кислорода по градиенту от низкой концентрации О2 к более высокой. Наивысшая концентрация кислорода по краям покровного стекла, где кислород диффундирует в воду из воздуха, и в непосредственной близости от нити водоросли, где кислород выделяется как побочный продукт фотосинтеза.
в) Можно оставить препарат в темноте примерно на 30 мин и снова рассмотреть его. Все бактерии должны теперь скопиться у краев покровного стекла, так как водоросль в темноте не фотосинтезирует.
15.6. а) Световой стимул воспринимается кончиком колеоптиля. Затем какой-то сигнал передается от кончика (рецептора) в область ниже кончика (эффектор).
6) Опыт В был нужен для проверки результатов опыта Б, которые могли быть следствием повреждения колеоптиля.
15.7. Были получены дальнейшие данные о существовании какого-то сигнала - по-видимому, химического вещества (гормона). Это вещество не может пройти через непроницаемый барьер. Оно передвигается главным образом вниз по затененной стороне колеоптиля. В опыте Б слюда мешает такому передвижению. Поэтому свет либо ингибирует образование гормона, либо вызывает его инактивацию (стимулирует его распад), либо заставляет его перераспределяться в латеральном направлении.
15.8. См. рис. 15.8 (отв.).
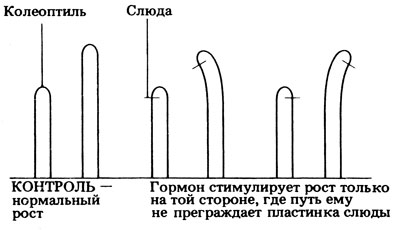
Рис. 15.8 (отв.). Повторение опытов Бойсен-Йенсена при равномерном освещении. Показаны три эксперимента; во всех случаях слева представлены условия опыта, справа - результат
15.9. Кончик колеоптиля образует какое-то химическое вещество, которое диффундирует в агар. Оно может стимулировать рост в зоне, расположенной ниже кончика, и восстанавливает нормальный рост (опыт А). В условиях равномерного освещения или в темноте практически не происходит никакой латеральной передачи этого вещества (опыт Б).
15.10. Колеоптиль должен будет расти в левую сторону.
15.11. А-100 ч. на млн.; Б-10 ч. на млн.; В-1 ч. на млн.; Г-0,1 ч. на млн.; Д-0,01 ч. на млн.; Е-нуль.
15.12. Относительно высокая концентрация ауксина стимулирует рост колеоптилей (или побегов), но ингибирует рост корней. Это подтверждает выводы, сделанные из опыта 15.1.
15.13. См. разд. 16.5.6.
15.14. а) Абсцизовая кислота может транспортироваться из кончиков корней вверх, претерпевать латеральный транспорт в тканях корня в ответ на воздействие силы тяжести и подавлять рост.
б) ИУК, вероятно, не участвует в геотропической реакции корня кукурузы, поскольку она, по-видимому, не транспортируется из кончика корня вверх.
15.15. а) крахмал;
б) мальтоза;
в) мальтаза.
г) Крахмал, запасенный в эндосперме, служит главным питательным веществом семян хлебных злаков.
15.16. Запасные белки перевариваются (гидролизуются) с освобождением аминокислот - основных структурных единиц белка. Из этих аминокислот синтезируются ферменты (все ферменты - белки), например α-амилаза; эти ферменты используются затем для переваривания питательных веществ эндосперма.
15.17. Амилазная активность может быть связана с присутствием микроорганизмов на коже рук или же с попаданием слюны на пальцы изо рта. Поэтому в подобного рода опытах очень важно не брать в руки семена, после того как их поверхность простерилизована.
15.18. Можно инкубировать семена с радиоактивными (мечеными 14С) аминокислотами. Это приведет к образованию меченой амилазы. Можно также инкубировать семена с ингибиторами белкового синтеза (например, с циклогексимидом); это помешает синтезу амилазы, и тогда не будет обнаружено никакой амилазной активности.
15.19. Разделив семена на алейроновую и неалейроновую части, можно показать, что меченая амилаза появляется сначала в алейроновом слое. Можно также раздельно инкубировать эндосперм с алейроновым слоем и эндосперм без этого слоя на агаре с крахмалом и гиббереллином; тогда амилаза должна синтезироваться только в первом случае (практически, однако, достичь такого результата нелегко).
15.20. Одна из лучших биологических проб на гиббереллин (быстрая, надежная и чувствительная) основана на инкубации зародышевых половинок семян ячменя с исследуемым материалом. Через два дня содержание редуцирующих Сахаров в зародышах будет пропорционально количеству гиббереллина в этом материале.
15.21. а) В молодом листе аминокислота задерживается и не уходит далеко от места ее нанесения. В старом листе часть ее экспортируется по жилкам и черешку.
б) Молодой лист использует аминокислоту в процессе роста для построения белков. Старый же лист уже не растет и поэтому экспортирует питательные вещества в другие части растения, например в корни и молодые листья.
в) Ткань, обработанная кинетином, удерживает аминокислоты или даже "притягивает" их. (Причины этого неизвестны; возможно, это связано с тем, что кинетин либо поддерживает, либо стимулирует нормальную активность клетки.)
15.22. Можно, например, взять такое растение, о котором известно, что добавленный гиббереллин влияет на рост его стебля, и удалить собственный источник ауксинов, отрезав верхушку стебля. Тогда гиббереллин должен оказаться неэффективным. Очень важно показать, что ответную реакцию растения можно восстановить, добавив ауксин (например, ИУК в ланолиновой пасте), так как отсутствие реакции на гиббереллины могло бы быть обусловлено повреждением растения или же влиянием какого-то другого фактора. Такие опыты действительно демонстрируют полную зависимость реакции от ауксина.
15.23. а) Ауксин (ИУК). б) См. рис. 15.23 (отв.).
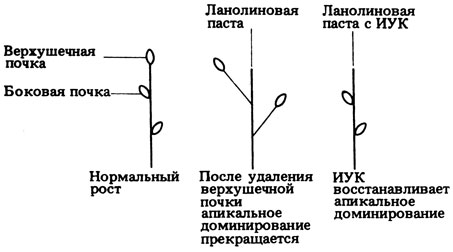
Рис. 15.23 (отв.). Эксперимент, показывающий роль ИУК в доминировании верхушки
15.24. Мелкие листья легче проходят сквозь почву (листья злаков остаются внутри колеоптиля). Изогнутые крючком почечки зародыша у двудольных растений предохраняют нежную верхушечную меристему от повреждения частицами почвы. Удлиненные междоузлия дают листьям максимальные шансы выйти на поверхность, к свету.
15.25. См. гл. 9.
15.26. График представлен на рис. 15.26 (отв.). Он демонстрирует противоположное действие красного и дальнего красного света. 30-секундная экспозиция на красном свету (при интенсивности освещения, использованной в данном эксперименте) полностью аннулирует индуцирующее действие длинной ночи. Эффективность красного света возрастает с увеличением времени экспозиции до 30 с. Эффект красного света можно полностью снять дальним красным светом, хотя для этого нужна более длительная экспозиция (50 с). Эти результаты позволяют предполагать, что фоторецептором служит фитохром.
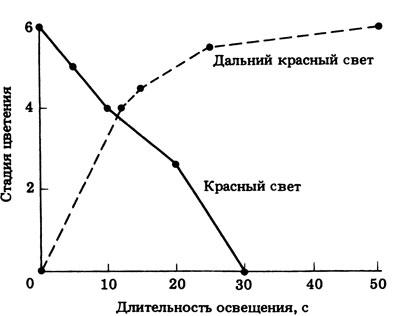
Рис. 15.26 (отв). Влияние прерывания 'длинной' ночи красным и дальним красным светом на цветение дурнишника
15.27. Можно использовать несколько методов. Один из простейших опытов показан на рис. 15.27 (отв.). Прямоугольниками обведены части растения, укрываемые от света для создания эффекта короткого дня.
15.28. Подавление роста пазушных почек, или доминирование верхушки, обусловлено в основном действием ауксинов. (О доминировании верхушки см. в разд. 15.3.3).
|
ПОИСК:
|
При копировании ссылка обязательна:
http://biologylib.ru/ 'Библиотека по биологии'